1)Fusobacterium nucleatum(Fn)
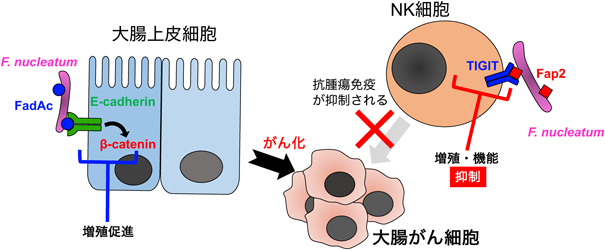
もともと口腔内の歯周病菌として知られていたFusobacterium nucleatumが大腸がん患者の腫瘍部付近に正常組織周囲よりも多く検出されることが明らかになり,その発がん促進性について研究が進んでいる.F. nucleatumは,大腸がん患者の腸内細菌叢解析の複数の大規模コホートで常に上位に位置しており3–5),大腸がんの発症や進展に寄与していると考えられている.実験的には,腸の腫瘍を発症しやすいApcMin/+マウスに炎症性腸疾患患者由来のF. nucleatum種を移植すると,ApcMin/+マウスの腸の腫瘍数が有意に増加し,そのような状況では周囲の微小環境が変化し,骨髄系の細胞が有意に増加していることが示された6).さらに,F. nucleatumの腫瘍原性についても分子メカニズムが示され,菌体表面に発現するFadA adhesion protein complex(FadAc)が,大腸上皮の細胞の細胞接着因子,Eカドヘリン(E-cadherin)に結合しβカテニン(β-catenin)シグナルを活性化し,細胞増殖を促進することが報告されている(図1)7).さらに最近の報告では,F. nucleatumはこのような増殖促進性だけでなく,抗腫瘍免疫の抑制にも働くことが報告された.ヒトのNK(natural killer)細胞の抑制性受容体TIGIT(T cell immunoglobulin and ITIM domain)にF. nucleatumがFap2を介して直接結合し,抗腫瘍免疫を抑制し,大腸がんの促進に関与することが示唆された(図1)8).F. nucleatumの数が多い患者の方が大腸がんの予後が悪いという結果もあり,本菌は大腸がんの進展や予後予測に使える可能性が示唆されている.
F. nucleatumは,大腸がんの発症や進展に寄与することが示されているが,筆者らはもともと口腔細菌であるF. nucleatumが,口腔がんの進展に関係するのかどうか明らかにするため,口腔がん患者の唾液,歯垢,腫瘍部のバイオプシーサンプルを収集し,菌叢を調べた9).その結果,やはりF. nucleatumが,口腔がん腫瘍部のサンプルで最も多く検出された.F. nucleatumは五つの亜種の存在が知られているが,筆者らが調べた患者サンプルでは,Fn. polymorphumが多く検出された.患者の唾液サンプルから単離したFn. polymorphumを用いて,舌がんを発症しやすい遺伝子改変マウスモデル10)に菌を塗布したところ,コントロールマウスと比べて,有意な舌上皮細胞の増殖とYAP(Yes-associated protein)の活性化が認められた.YAPの活性化はヒト舌がんで高頻度に認められることが知られており,F. nucleatumがYAPの活性化を促進する可能性が示唆された9).
2)変異原性毒素(genotoxin)産生菌
a.pks陽性Escherichia coli
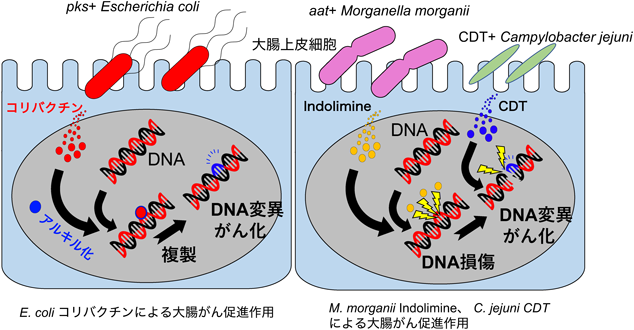
ある種の大腸菌(Escherichia coli)はpolyketide synthase(PKS)をコードする遺伝子pksを持ち,コリバクチン(Colibactin)と呼ばれる強い変異原性を持つ毒素(genotoxin)を産生するが,これが大腸がんの原因となることが示された11).この報告では大腸炎を発症しやすいIL-10ノックアウトマウスを用いて,アゾキシメタンを用いた大腸がん誘発化学発がん実験を行い,どのような腸内細菌が有意に増加するかを調べた.増加した菌のうち,E.coli(NC101株)が悪性度の強い大腸がんの発症を促進することが明らかになった11).この報告から,腸炎とpks陽性のE.coliが相乗的に大腸がんを促進する可能性があることが示唆された.pks陽性のE.coliが産生するコリバクチンのDNAに対する作用について,さらに検討がなされ,驚くべきことに,コリバクチンはDNAをアルキル化し,コリバクチンのDNAへの付加体を形成することが明らかになった12)(図2).このDNA付加体は次の複製時にDNA変異を起こすため,この論文の著者らはコリバクチンが付加されたDNAが大腸がんの危険因子としてのバイオマーカーになりうると述べている12).コリバクチンが関与する変異は特徴的なシグネチャーを示し,大腸がんの発症に寄与するとともに,診断に利用できる可能性がある13).
b.aat陽性Morganella morganii
腸内細菌が産生する新たなgenotoxinとしてIndolimineが報告された(図2).Indolimineは,ロイシン,バリン,フェニルアラニンを脱炭素酵素により代謝し,indole-3-aldehydeと結合することで合成される新規腸内細菌代謝物である.Caoらは,潰瘍性大腸炎患者から分離した122株の腸内細菌の代謝物からDNAを直接損傷する分子をスクリーニングし,Morganella morganiiが産生する小分子が直接DNAを切断することを突き止めた.質量分析装置とNMRを用いて分子の構造を決定し,新規の分子としてIndolimineと名づけた.そして,M. morganiiの全ゲノムシーケンスからIndolimine合成にはaspartate aminotransferase(AAT)が必要であることを見いだした.aat陽性のM. morganiiは大腸がん誘導モデルマウスにて腫瘍形成数を増加させることから,M. morganiiが産生するIndolimineが新たなgenotoxinであると結論づけた14).
c.cytolethal distending toxin(CDT)産生Campylobacter jejuni
Campylobacter jejuniもcytolethal distending toxin(CDT)というgenotoxinを産生することが知られている.CDTはDNAse活性があり,DNA二本鎖損傷を促す(図2).無菌のApcMin/+マウスにヒト由来のC. jejuniを定着させた研究で,腸の腫瘍が促進されたという報告がある15).また,C. jejuni由来のCDTを含むメンブレンベシクルが,宿主の細胞に結合し,細胞増殖を止める作用があることが報告された16).
3)enterotoxigenic B. fragilis(ETBF)
難治性の潰瘍性大腸炎患者の数%が大腸がんを併発することが知られており,腸内細菌が関わる腸管の炎症が大腸がんの発症を促進するという研究結果が報告されている.その一つはBacteroides fragilis(B. fragilis)を介する大腸がんである17).B. fragilisにはB. fragilis toxin(BFT)という亜鉛依存性metalloproteaseを分泌するenterotoxigenic B. fragilis(ETBF)とBFTを分泌しないnon-toxigenic B. fragilis(NTBF)が存在する.Searsらのグループは,腸に腫瘍を形成しやすいApcMin/+マウスを用いて,ETBFを腸に移植すると腸炎や大腸がんが発症するのに対し,NTBFを腸に移植したマウスは腸炎も大腸がんも発症しないことを見いだした.そのメカニズムとして,SearsらはETBFの産生するBFTが,腸内のTh17細胞を刺激し,Stat3を活性化することが炎症や腫瘍発生の原因であることを示した18).マウスモデルを用いたこれらの知見がヒトにおいても示されている.すなわちヒトの大腸アデノーマにおいて,ETBFがバイオフィルムを形成しており,炎症を持続的に惹起し,大腸発がんに寄与していることが示された19).また大腸がん患者で増えていることがよく知られている,F. nucleatumも菌の産生する短鎖脂肪酸を介してTh17の活性化に寄与することが示され20),したがってB. fragilisとF. nucleatumが炎症性大腸がんの発症に寄与している可能性がある.
4)Streptococcus gallolyticus subspecies gallolyticus(Sgg)(Streptococcus bovis biotype Iより名称変更)
古くから,Streptococcus gallolyticus subsp. gallolyticus(Sgg)による感染性心内膜炎患者で大腸がんを合併する症例が多い(47~85%)ことが報告されていたが21),近年多くの菌叢解析から,Sggが大腸がんと関係していることは確かな現象と認識されている.また,大腸がん患者の68%で,血中のSgg特異的抗体が陽性になることから,大腸がんのリスク診断の一つの指標ともなっている.Sggは大腸がんの組織に定着しやすく,腫瘍血管から血中に侵入し,菌血症の頻度が増加するため,感染性心内膜炎へと進展する可能性が高くなるのではないかと推察されている.このようにSggは大腸がんの発症と強い相関があるものの,大腸がんを発症させるドライバーとしては考えられていないようである.Sggは大腸がんの組織に定着しやすい性質があるが,Sgg特異的なgallocinという毒素を産生し22),周囲の菌を殺菌して自身が増える微小環境を形成するため,大腸がん患者で高い相関性をもって存在すると考えられている.
1)腸管バリアの脆弱性
近年,腸内細菌がさまざまな肝疾患に影響を及ぼすことがわかってきた.その原因として,飲酒や長期にわたる高脂肪食摂取,また,ある種の腸内細菌により生じる腸管バリアの脆弱化があげられる.腸管バリアの脆弱化が生じると,腸内細菌由来物質が肝臓に移行し,肝臓においてMAMPsやPAMPs(microbial- or pathogen-associated molecular patterns)による炎症シグナルが活性化され,肝臓における慢性炎症や肝細胞がんの発症に寄与する可能性がある.実際にヒトの脂肪肝や脂肪性肝炎の患者では,リポ多糖類(lipopolysaccharide:LPS)を供給する大腸菌が腸で増加しており,このような腸内バリアの機能不全により,門脈を介した腸内細菌由来物質の肝臓への蓄積だけでなく,肝臓を経由して腸内細菌由来物質が全身性に増加することが知られている.また,菌体成分だけでなく,腸内細菌の産生する代謝物質が,サイトカイン・ケモカイン等の産生を介して,肝組織微小環境を変化させ,肝疾患へ影響する場合があることもわかってきた.本節では,腸内細菌叢が関わる肝がんについて,筆者らの研究内容および,最近報告された興味ある知見を紹介する.
2)胆汁酸の影響
コール酸(cholic acid:CA)やケノデオキシコール酸(chenodeoxycholic acid:CDCA)などの一次胆汁酸は,宿主の肝臓の酵素によってコレステロールから産生されるが,CAとCDCAからそれぞれ形成されるデオキシコール酸(deoxycholic acid:DCA)やリトコール酸(lithocholic acid:LCA)などの二次胆汁酸は,7-α-脱ヒドロキシル化を行う胆汁酸誘導性オペロン(bile acid inducible operon:bai operon)を持つ腸内細菌叢によって産生される.また,CDCAなどのいくつかの胆汁酸は,FXRなどの核内受容体のリガンドとして作用する.FXRを介したシグナル伝達の刺激は,胆汁酸産生経路の酵素の発現を変化させ,胆汁酸の産生量を調節する.FXRによるシグナルは肝障害モデルにおける脂肪性肝炎,門脈圧亢進症,および肝炎症を改善する可能性があることが報告されており,実際,FXRアゴニストは有望な抗肝炎薬候補となっている23).また,後述するが,特殊な修飾を受けたリトコール酸3-oxoLCAとisoLCAは,Th17細胞のマスター転写因子であるRORγtへの直接結合を介して,その転写活性を抑制し,Th17細胞への分化を阻害することが示されている24).このように,核内受容体に結合した胆汁酸が媒介する転写シグナルは,肝疾患に大きな影響を与える可能性がある.
3)腸内細菌と脂肪性肝炎関連肝がん
脂肪性肝疾患を素地とする肝がんには,肝硬変を経由するものと,肝硬変をほとんど認めず発症にいたるものが約半数ずつ存在する.ウイルス性肝炎にともなう肝がんは肝硬変を伴うものがほとんどであるため,肝硬変を伴わない脂肪性肝疾患関連肝がんには,それ特有の発症機構が存在する可能性がある.
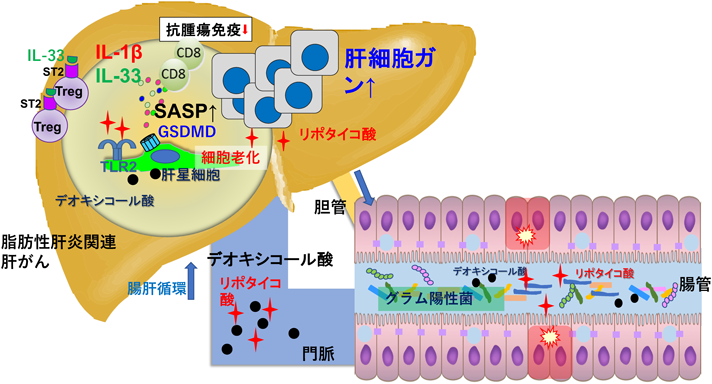
筆者らは新生仔マウスにDMBA(7,12-dimethylbenz[a]anthracene)を1回のみ塗布し,その後群を分け,高脂肪食または普通食を摂取させたところ,DMBAを塗布し普通食を摂取させたマウスに比べて,DMBAを塗布し高脂肪食を摂取させたマウスの方が有意に肝がんを多く発症することを見いだした25).さらに肝がん組織の詳細を調べたところ,肝星細胞において細胞老化が生じており,細胞老化に伴って発がん促進作用のある多くの炎症性サイトカインやプロテアーゼなどが分泌される細胞老化随伴分泌現象(senescence-associated secretory phenotype:SASP)と呼ばれる現象が生じ,発がん促進的な微小環境を形成していることが明らかになった.このモデルマウスでは高脂肪食摂取で増加するグラム陽性細菌がDCAを産生しており,腸肝循環するDCAが肝星細胞の細胞老化を誘導することがわかった.
また,この肝発がんモデルマウスの肝臓の類洞領域にはリポタイコ酸(lipoteichoic acid:LTA)が著しく蓄積しており,抗生剤投与でその蓄積が減少することから,このLTA蓄積は高脂肪食摂取によって増加したグラム陽性腸内細菌の成分が肝移行したことによることがわかった.また,肝臓微小環境におけるDCAとLTAの増加は,プロスタグランジン産生の律速酵素であるシクロオキシゲナーゼ2(cyclooxygenase-2:COX-2)の発現,ならびにDCAにより細胞老化を起こした肝星細胞におけるSASP現象を助長し,SASP因子の産生を相乗的に増加させることがわかった.COX-2によるプロスタグランジンE2(PGE2)産生は抗腫瘍免疫を抑制しており,肝がんの進展を促進している可能性も示唆された.COX-2の過剰発現とPGE2の過剰産生はヒト脂肪性肝炎関連肝がんでも観察され,このメカニズムがヒトの肝がんの進展にも関与している可能性があることが示唆された26).
さらに最近筆者らは,脂肪肝に蓄積したLTAがトリガーとなり,肝星細胞でガスダーミンDが切断されN末端の切断体が検出されることを見いだした.ガスダーミンDはがん微小環境の細胞老化した肝星細胞で高発現しており,SASP因子のIL-33とIL-1βが,切断されたガスダーミンDのN末端により形成される小孔から放出されることが明らかになった.ガスダーミンDのN末端切断体による小孔が形成されると,パイロトーシスという細胞死の一種が促進されることが知られている.しかし,老化した肝星細胞では,細胞膜上で小孔が形成されても,パイロトーシス細胞死は非常に生じにくく,パイロトーシスに抵抗性が生じていることが明らかになった.老化細胞でパイロトーシスが生じにくいことは,老化細胞からのSASP因子の放出が続くことを示唆しており,SASP現象が持続するメカニズムの一つと考えられる.放出されたIL-33は,その受容体ST2(Serum Stimulation-2,別名Il1rl1とも呼ばれる)を発現する制御性T細胞(regulatory T Cells:Treg)細胞を活性化し,その結果,抗腫瘍免疫が抑制され,肝がんが進行していくことが明らかになった27)(図3).
ヒトの脂肪性肝炎関連肝がんの腫瘍部に存在する肝星細胞において,ガスダーミンDのN末端切断体の特異的抗体を用いて調べたところ,確かに切断体が認められた.このことから,マウスモデルで検証したこれらの知見は,ヒトの脂肪性肝炎関連肝がんの一部においても同様に働いている可能性が示唆された.これらの結果から,ガスダーミンDによる小孔形成の阻害や,放出されたIL-33が作用するST2を発現するTreg細胞の機能阻害は,肝がんの予防や治療の標的として使用できる可能性がある.
4)一次胆汁酸による肝類洞内皮細胞を介したNKT細胞による抗腫瘍免疫の活性化
Gretenらのグループは,一次胆汁酸で刺激された肝類洞内皮細胞からのケモカインを介する抗腫瘍免疫の活性化と肝がん形成制御に関する研究を報告した28).Mycを活性化する肝がん発症のマウスモデルで,抗生剤を投与しておくと,肝がんの発症が抑えられる.次に彼らはその機構として,抗生剤投与により,抗腫瘍免疫を担う肝臓のNKT細胞が活性化することを見いだし,NKT細胞活性化のメカニズムとして,胆汁酸に着目した.抗生剤投与により,体内の二次胆汁酸は減少するが,多くの一次胆汁酸は逆に増加する.抗生剤投与により増えた一次胆汁酸,特にタウロベータ-ムリコール酸(tauro-β-muricholic acid)により,肝類洞内皮細胞からCXCL16が産生されるが,逆に,DCAなどの二次胆汁酸は,肝類洞内皮細胞からのCXCL16産生を抑制する.CXCL16はその特異的受容体であるCXCR6を発現するNKT細胞に作用し,この細胞を強く活性化して,NKT細胞による抗腫瘍免疫が発揮されることが明らかになった.本研究により,一次胆汁酸が肝類洞内皮細胞へ作用し,肝臓におけるNKT細胞を活性化し,肝がんの進展を抑制している機構が明らかになった28).
5)腸内細菌とSVR後肝がん
近年,肝炎ウイルスに対する治療薬の開発が進み,特にC型肝炎ウイルスは,抗ウイルス薬の内服により除去できる時代になった.ウイルスが消失した状態を,ウイルス学的持続陰性化(sustained virologic response:SVR)という.しかし,ウイルスを除去しても肝がんが発症・進行する症例が10%程度存在し,このような症例では,どのようなメカニズムで肝がんが発症するのか,解明が待たれる.ウイルス感染によって変化した,肝実質細胞や肝臓における免疫状態や肝線維化が,後の肝がん発症に関係するとの予測がされているが,いまだ,明確なメカニズムは解明されていない.最近,SVR後肝がんの発症に腸内細菌が関与している可能性があるという研究結果が報告された29).C型肝炎患者では,Enterococcus faecalisが腸内に多いことを見いだし,C型慢性肝炎患者,C型肝炎背景の肝がん患者の便を移植した,肝線維化背景の肝がんモデルマウスでは肝がんが有意に多く発症することを確認した.詳細を調べた結果,E. faecalisの中でも,プロテアーゼを発現するgelE陽性E. faecalisが腸管バリアを脆弱化し,LPSによりTLR4シグナルが活性化し,肝がんの進展に関わる可能性を示した.前項に記載した非ウイルス性の脂肪性肝炎関連肝がんと違って,二次胆汁酸のDCAの患者血中濃度は低いことが示され,二次胆汁酸産生菌の減少が,gelE-positive E. faecalisの増加につながっていることが示唆された.上記した脂肪性肝炎関連肝がんで,二次胆汁酸のDCAの増加が発がんに関係する現象とは逆である.SVR後肝がんの発症に特定の腸内細菌種が関係する可能性があるという,非常に興味深い結果である.
6)LPSは線維化背景の肝細胞がん,胆管がんの発症に関与する
腸管バリアの脆弱化によるLPSの肝移行は,肝臓において持続的な低レベルの炎症を起こす.Schwabeらのグループは,LPS/TLR4による持続的な炎症シグナルが,diethylnitrosamine(DEN)および四塩化炭素投与によるマウス肝がんモデルにおいて,肝がんを促進することを報告した30).また最近,原発性硬化性胆管炎(primary sclerosing cholangitis:PSC)に伴う胆管がんの進行もグラム陰性菌由来のLPSによって加速されることが報告され,そのメカニズムとしてLPSがTLR4(Toll-like receptor 4)を介して骨髄由来免疫抑制細胞(myeloid derived suppressor cells:MDSC)を活性化することが報告された31).胆管結紮によるPSCモデルマウスの腸内では,Verrucomicrobiales目などのグラム陰性菌が増え,回腸を中心に著しい炎症が生じていた.また,胆管結紮によるPSCモデルマウスに生じた胆管がんでは,腫瘍部にCXCR2陽性のMDSCの蓄積を認めた.腸管バリアの機能不全はPSC患者でも観察される.Klebsiella pneumoniaeなどPSC患者で多く認められる腸内細菌は,腸上皮を傷害する能力が強いことが別の研究で示されていたが,PSC患者に多い腸内細菌により腸管バリアが脆弱化すると,肝臓中に腸内細菌叢由来のLPSが増加すると考えられる.増加したLPSは,TLR4依存的に肝細胞においてCXCL1発現を誘導し,次いでCXCL1は肝臓において,MDSCのサブセットの一つで抑制性の免疫細胞,PMN(polymorphonuclear)-MDSCをリクルートする.PMN-MDSCは,胆管がん微小環境における抗腫瘍免疫を抑制し,胆管がんを進行させる.この研究ではネオマイシン処理により,CXCL1発現とPMN-MDSCの蓄積が抑制され,その結果,腫瘍増殖も抑制された.これらの結果から,腸内細菌叢由来のLPSが,胆管がんの進展に寄与していることが示された.
7)短鎖脂肪酸による肝がんの促進
短鎖脂肪酸(特に酪酸)は,腸管においてエピジェネティックなメカニズムにより,Foxp3というTreg細胞のマスター転写因子の発現誘導を介してTreg細胞への分化を促し,腸の炎症を抑制することが示されている32).腸内細菌の代謝物である短鎖脂肪酸は,抗炎症や肥満の予防に役立つことが多くの研究で示唆されている33–35).一方で,最近の研究では,イヌリンなどの水溶性食物繊維を過剰に摂取し,それにより短鎖脂肪酸のような発酵性代謝物が過剰に産生されると,胆汁うっ滞が生じ,二次胆汁酸が増加し,最終的には肝がんが促進されることが示された36).興味深いことに,セルロースなどの非水溶性食物繊維では胆汁うっ滞は生じていなかった.この胆汁うっ滞や肝がんの発症は腸内細菌叢に依存した病態であり,抗生剤投与マウスや無菌マウスでは生じなかった.肝がんに至る原因として,胆汁うっ滞による肝実質細胞のダメージ,それに伴う好中球の浸潤などが考えられた.発酵腸内細菌の除去により短鎖脂肪酸量が減少し,これらの病態は改善され,肝がん発症も抑制された.胆汁酸の吸収阻害薬も効果的であった.これらのことから,過剰に産生された短鎖脂肪酸が病態の増悪に関与したことが示唆された.この研究結果から,食物繊維の摂取も適度な量にとどめておくことが大切であると考えられる.
1)Th17細胞に影響する腸内細菌
Th17細胞は病原体の感染防御に重要なヘルパーT細胞であるが,一方でその過剰な反応は炎症性腸疾患(クローン病,潰瘍性大腸炎)やPSC,他の自己免疫疾患の病態にも深く関わっていることが近年明らかになっており,腸内細菌叢との関連が注目されている.
マウスの腸内常在細菌の一種であるセグメント細菌(segmented filamentous bacteria:SFB)は,多くの動物種で小腸,特に回腸末端に生息する菌種であることが知られている.SFBは粘液溶解遺伝子を持ち,粘液バリアを通過して上皮内層に付着することができるが,病原体とは異なり,上皮層に侵入することはない.また,SFBの上皮接着は宿主特異性を持ち,宿主以外の生物の腸管上皮には接着しないとされている37).興味深いことに,SFBはTh17細胞を誘導することが報告された37).SFBの接着はまた,血清アミロイドAタンパク質(serum amyloid A:SAA)の発現誘導とも関連している.SAAは腸管の粘膜固有層のCD11c+細胞を活性化してIL-1βを産生させ,それが腸管上皮細胞を刺激してより多くのSAAを分泌させるという,ポジティブ・フィードバックループを形成する.慶應義塾大学の本田らは,無菌マウスに健常人および潰瘍性大腸炎患者の糞便を定着させることでマウスの大腸粘膜にTh17が誘導されることを報告した.また,これらのマウスの腸内細菌を解析し,Th17の誘導に関わる20種のヒト腸内細菌を同定した.このことから,ヒトの腸管にもSFBと同様にTh17を誘導し腸管炎症を引き起こす腸内細菌が存在することが示された37).
また,慶應義塾大学の金井,中本らのグループは,難治性疾患であるPSC患者の便中に,肝臓内のTh17細胞の活性化を引き起こす3種類の腸内細菌,Klebsiella pneumoniae, Proteus mirabilis, Enterococcus gallinarumが高確率で存在することを,PSC患者の糞便を移植したモデルマウスの腸間膜リンパ節における菌解析から単離・同定した38).この3菌はいずれもPSC患者の便中に高い頻度で存在しており,特にK. pneumoniaeは18名の患者のうち17名で検出され,健常者,潰瘍性大腸炎,他の自己免疫性肝疾患の患者と比べて非常に高率であったことが報告されている.中でもK. pneumoniaeは大腸の上皮に穴を開け腸管バリアを破壊し,腸管の外にあるリンパ節に移行(バクテリアルトランスロケーション)し,肝臓内の過剰な免疫応答を誘導することが示された.さらに,同マウス肝臓で起こるTh17免疫反応は,抗菌薬によるクレブシエラ菌の排除により30%程度までに減弱することが示され,腸内細菌を標的とする治療法の可能性が示された38).
2)Treg細胞を誘導する腸内細菌
Treg細胞は,過剰な免疫反応を抑える機能を持つ抑制性免疫細胞であり,免疫寛容の成立や免疫細胞の恒常性の維持に必要である.Treg細胞は通常転写因子Foxp3が陽性であり,これがマスターレギュレーター転写因子でありマーカーとして使用される.腸など末梢に存在するpTreg(以前はiTregと呼ばれていた)と胸腺由来のtTreg細胞(HeliosおよびNeuropilin 1が陽性で,以前はnTregと呼ばれていた)に大きく分類される.Treg細胞は抑制性分子CTLA4を構成的に細胞表面に発現しており,IL-10, IL-35, TGF-βの産生を介して,抗原特異的T細胞を含めT細胞全般を阻害することができる.
慶應義塾大学の本田らのグループは,ヒト健常者の糞便の解析から,芽胞形成性クロストリジウム属のクラスターX IVaを含む17種の腸内細菌種によって,Treg細胞が誘導され,腸炎を抑制する可能性を報告した39).クロストリジウム属の中では,クロストリジウム・ラモサム(Clostridium ramosum)単独で大腸のTreg細胞を誘導できることが示されている40).
3)腸内細菌が産生する短鎖脂肪酸はTreg細胞を誘導する
上にも述べたように,クロストリジウム属の菌がTreg細胞を誘導する能力があることが明らかになったが,そのメカニズムは不明なままであった.この課題に対して理化学研究所の大野らのグループは,腸内細菌が産生する短鎖脂肪酸がTreg細胞を誘導することを報告した.多くの腸内細菌は食物繊維を短鎖脂肪酸に発酵させる能力がある.大野らは,腸炎のマウスモデルでは,短鎖脂肪酸の中でも特に酪酸がTreg細胞の数と相関性があったことに注目した.酪酸はヒストン脱アセチル化酵素阻害剤として作用し,Foxp3遺伝子の発現制御領域の非コード配列1(CNS1)に結合するヒストンのアセチル化を促進することによりオープンクロマチン化し,Foxp3の発現をエピジェネティックに増強する.その結果,Treg細胞が増加することが示された32).短鎖脂肪酸はまた,免疫細胞上のGPR43やGPR109aといったGタンパク質共役型受容体を介してシグナルを伝達し,Foxp3+ Tregの分化を促進し腸炎を抑制することが報告されている41, 42).
4)腸内細菌が産生する特殊な二次胆汁酸はTh17細胞への分化を抑制する
胆汁酸は宿主と腸内細菌叢,両方の代謝によって産生されるステロイド骨格を持つ化合物であり,一次胆汁酸は宿主の肝臓における複数の酵素反応により,コレステロールから産生される.肝臓ではヒトの場合,主にコール酸とケノデオキシコール酸の二つの一次胆汁酸が産生される.一方,二次胆汁酸は腸内細菌叢の代謝によって産生される.肝臓で抱合された修飾部位が脱抱合されたのち,7α-デヒドロキシラーゼを発現する菌属により,7α脱水酸化反応が起こり,二つの二次胆汁酸,3α-モノ水酸化胆汁酸[ケノデオキシコール酸から生じるLCA],そして3α-12α-ジ水酸化胆汁酸[コール酸から生じるDCA]が産生される.Clostridium scindensやClostridium hylemonaeなどのごく一部の腸内細菌は,完全な7α-脱水酸化経路の遺伝子(Baiオペロン)を持つことが知られる.さらに,いくつかの菌種は,Baiオペロンの有無にかかわらず,酸化反応やエピマー化反応を仲介して,オキソ(oxo),イソ(iso),アロ(allo)型の胆汁酸を生産することができる.たとえば,Eggerthella lentaは,3α-ヒドロキシステロイドデヒドロゲナーゼ(3αHSDH)の作用により,DCAとLCAからそれぞれ3-oxoDCAと3-oxoLCAを生成することができる.E. lentaはまた,3βHSDHの作用により,3-oxoDCAと3-oxoLCAをそれぞれisoDCAとisoLCAに変換することができる.Odoribacteraceae spp.やParabacteroides merdaeなどのバクテロイデス属は,二次胆汁酸からではなく,5α-レダクターゼと3βHSDHの作用により,3-oxo-4-LCAからisoalloLCAを合成する.非常に興味深いことに,これらの特殊な修飾を受けた二次胆汁酸は,Th17細胞の分化を制御することが報告された.特に3-oxoLCAとisoLCAは,RORγtへの直接結合を介して,その転写活性を抑制し,Th17細胞への分化を阻害することが示された24).ヒトのコホート研究においても,米国で実施された前向き研究(マサチューセッツ総合病院,PRISM)43)では,炎症性腸疾患(以下IBD)を認めるIBDコホート群とIBDを認めないコントロール群の糞便サンプルの比較では,3-oxoLCAとisoLCAのレベルが,IBDのクローン病患者で減少していることが確認された.さらに興味深いことに,Th17への分化を抑えるこれらの特殊なLCAは,ヒトの百寿者の腸内で多いことが報告された.このことは,腸炎を抑制するような二次胆汁酸が長寿と関係する可能性を示唆している44).
5)ポリアミンによるT細胞制御
ポリアミン(プトレシン,スペルミン,スペルミジン)は消化管内に豊富に存在し,T細胞の制御に関与しており,近年,抗腫瘍作用や抗老化作用が注目されている.ポリアミンは食品としての経口摂取するほか,生体内でもオルニチンから生合成されるが,腸内細菌による産生も多く腸内細菌は重要な供給源である.スペルミジンは,特定のリシン残基をヒプシン化する.ヒプシン化されたeIF5Aはミトコンドリア分子の発現を促進し,TCAサイクルに依存したエネルギー代謝を促進するが,ポリアミン-ヒプシン経路が欠如すると,ヒストンアセチル化の変化とエピジェネティックなリモデリングが広範囲に起こり,CD4 T細胞の分化が乱れ,マウスは腸炎を起こすことが報告された45).このことは,ポリアミンを使ったヒプシン化によるeIF5Aの活性化がCD4 T細胞の正しい分化に重要な役割を果たしていることを示している.
さらに,最近,スペルミジンが,ヘルパーT細胞だけでなく,CD8 T細胞の活性,すなわち抗腫瘍免疫の活性化(若返り)に寄与すると報告された46).この研究は,老齢マウスでは免疫チェックポイント阻害剤(Immune-check point inhibitors:ICI)の効果が低いが,スペルミジンの併用で,ICIの効果が劇的によくなるという発見から始まった.この研究では,スペルミジンにより,ミトコンドリアにおけるβ酸化が活性化され,疲弊T細胞が減少することが明らかになった.さらに詳細を調べると,β酸化の律速酵素がスペルミジンによりヒプシン化し,活性化することが新規に明らかになった.このように,スペルミジンは,T細胞の活性を制御することが多く示されている.
6)腸内細菌とIgA
腸管の免疫細胞は,パイエル板などの腸管関連リンパ組織(gut associated lymphoid tissue:GALT)や,腸管粘膜固有層,および腸管上皮などに分布し,それぞれ存在する種類や機能が異なっている.パイエル板はリンパ組織であり,抗原に曝露されていないナイーブT細胞やB細胞が多く存在する.組織学的にはリンパ濾胞と濾胞間領域などの構造を有し,リンパ濾胞では胚中心においてB細胞がIgM産生性のものからIgA産生性のものへクラススイッチされる.腸におけるT細胞依存性のIgA誘導は,微生物叢を形成する重要なメカニズムの一つである.腸粘膜で微生物に曝露されると,IgA陽性形質細胞が生成される.IgA陽性形質細胞は反応性に優れ,各細菌の表面抗原に特異的なIgAが形成される.
理化学研究所の大野らのグループは,短鎖脂肪酸のうち,酢酸を付加したセルロースにより,IgA産生細胞やIgA分泌量が増加すること,また,IgAの腸内細菌に対する結合率も増加していることを見いだした.酢酸セルロース投与マウスのIgAはEnterobacterales目に属する大腸菌など病原性細菌に結合する傾向があることがわかった.この酢酸による病原菌特異的IgA誘導は,T細胞依存的であることが明らかになった47).
また,最近,大阪大学の原らのグループは,加齢で腸内細菌叢が変化するメカニズムをIgAの観点から明らかにした48).腸の胚中心B細胞は,常在細菌に継続的に曝露されることにより,加齢に伴い腸の細胞老化が生じ,これにより腸内細菌を標的とするIgA抗体の産生量と多様性の両方が減少し,その結果,加齢マウスにおける腸内細菌叢の組成が変化することをつきとめた.この研究結果から,腸内細菌叢と胚中心B細胞の間にIgAを介したクロストークが存在することが明らかになった.
最後に,最近特に注目されている腸内細菌と抗腫瘍免疫の関連について述べる.先行的に主に悪性黒色腫で研究されているが,今後,さまざまながんでこの機構が確認され利用される可能性がある.抗腫瘍効果を活性化する免疫チェックポイント阻害剤(ICI)として,免疫抑制性分子PD-1やそのリガンドPD-L1,またCTLA4などに対する抗体が,実臨床でも使用されている.しかし,すべてのがん症例でこのICIの効果があるわけではなく,また,irAE(immune-related adverse events)と呼ばれる重篤な免疫関連副作用にも気をつけなければならない.最近,ある種の腸内細菌が,抗腫瘍免疫力を高め,これらのICIの働きを助けるように作用することが報告された.悪性黒色腫の患者で免疫チェックポイント阻害剤の効果があった悪性黒色腫の患者では,Ruminococaceae属が有意に増えており49),大腸がん,非小細胞肺がんなど,上皮性のがんではAkkermansia種の腸内細菌が増えていることがわかった50).無菌マウスに,ICIが有効であった悪性黒色腫患者由来の腸内細菌を移植[糞便移植(fecal microbiota transplantation:FMT)]すると,抗腫瘍免疫が活性化し,有意に良好な抗腫瘍効果が観察された49).さらに最近,二つの臨床試験で,抗PD-1抗体抵抗性悪性黒色腫患者において,ICIが有効であった悪性黒色腫患者由来のFMTに加えて,ICIの再投与の安全性と実現可能性が評価された.ある研究では,10人の患者のうち3人51),また別の研究では,15人の患者のうち6人に有効性がみられた52).特に,FMTによる治療は,CD8+ T細胞活性化の増加を含む抗腫瘍性変化をもたらした.これらの結果は,ICIに対する耐性が生じても腸内微生物叢を変えることによって,それを克服できることを示唆している.このような現象が起きるメカニズムであるが,最近の研究では,FMTがICIに対する応答に影響を与えるのは,おそらく腸内細菌がアジュバントとして機能するためではないかと考えられている.ある種のエンテロコッカスのムロペプチド(リポタイコ酸の一種)がICIの効果を高めることが示唆されている53).
さらに,最近,非小細胞肺がん患者において,Akkermansia muciniphilaを腸内に有する患者の方がICIの効果が高いことや54),腎臓がん患者において細菌製剤のサプリメントがICIの効果を上げることが次々と報告されている55).今後,詳細なメカニズムを解明し,どのような症例でどのような菌または菌の成分がICIの効果を上げるのか明確にしていけば,実臨床のプロトコールにプロバイオティクス製剤として菌のサプリメント等が加わる可能性は十分にあると考える.また,アマゾン地方のカムカムフルーツと呼ばれる果物に含まれる天然化合物カスタラジンが腸内細菌叢を変え,ICIの効果を上昇させるプレバイオティックス食品として報告された.つまり,もともと,ICIに対して応答しないノンレスポンダー型の腸内細菌叢を持っていても,カスタラジンなどのプレバイオティックス食品を摂取すれば,レスポンダー型の腸内細菌叢に変化させることが可能となる56).今後,同様の効果を示すプレバイオティックス化合物のスクリーニングも活発化しそうである