セラミドとregulated cell deathCeramide and regulated cell death
摂南大学薬学部Faculty of Pharmaceutical Sciences, Setsunan University ◇ 〒573–0101 大阪府枚方市長尾峠町45–1 ◇ 45–1 Nagaotouge-Cho, Hirakata, Osaka 573–0101, Japan
発行日:2024年8月25日Published: August 25, 2024
セラミドはアポトーシスの生理活性脂質であり,長年にわたりその全容解明に向けた研究が展開されている.近年,アポトーシス以外に多様なプログラム化された細胞死が同定され,これらの細胞死はregulated cell death(RCD)として呼ばれており,これらRCDへのセラミドの関与が指摘され始めている.セラミドには多数の分子種があり,個々のRCDへのセラミド分子種特異的な関与を示す報告が蓄積しており,セラミド分子種の多様性が注目されている.本稿では,セラミド分子種に着目して,RCDの制御機構におけるセラミド生物学についてまとめた.
© 2024 公益社団法人日本生化学会© 2024 The Japanese Biochemical Society
セラミドは細胞死を制御する脂質として広く知られ,その全容解明に向けた研究が長年にわたり展開されている.1993年,Obeidらは細胞膜透過性を有する短鎖C2:0-セラミド(図1;人工合成されたセラミド)がアポトーシスを誘導することを報告した1).これが細胞死に関するセラミド研究の幕明けである.これ以降,細胞死制御へのセラミドの関与を示す研究報告が蓄積され,さらにはセラミドの生成・代謝酵素,輸送タンパク質や標的タンパク質の同定,制御機構の解明,スフィンゴ脂質質量分析技術の発展および創薬応用も急速に進んでいる2–7).
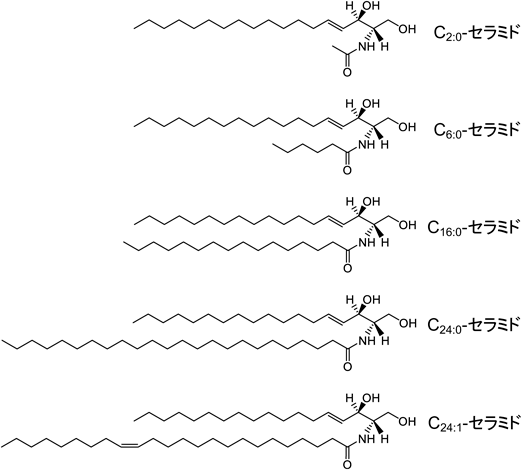
セラミドの側鎖には短鎖(炭素数10以下),長鎖(炭素数11–20)や極長鎖(炭素数21以上)の脂肪酸が結合している.C2:0-セラミド(短鎖セラミド)およびC6:0-セラミド(短鎖セラミド)は人工的に合成されたセラミド分子種である.天然には多様なセラミド分子種が存在しており,ここでは主要なセラミド分子種の構造を示す:C16:0-セラミド(長鎖セラミド),C24:0-セラミド(極長鎖セラミド)およびC24:1-セラミド(極長鎖セラミド).
20世紀では細胞死はネクローシスとアポトーシスに大別されており,前者は非プログラム化細胞死,後者はプログラム化細胞死として認識されていた.その後,ネクローシスまたはアポトーシス様形態を示す細胞死の中に,薬理学的もしくは遺伝的介入により制御できる新たな細胞死が多数発見された.これらの研究成果を整理するため,2018年,細胞死研究に携わる何百人もの科学者が共同で「Molecular mechanisms of cell death:recommendations of the Nomenclature Committee on Cell Death 2018」と題する論文をCell Death & Differentiation誌に掲載し,各細胞死の命名の統一が提案された8).そこでは,実質的・瞬間的で制御不能な細胞死を示すaccidental cell deathと一つ以上のシグナル伝達モジュールの活性化から生じる細胞死を示すregulated cell death(RCD)に分類され,外因性アポトーシス(extrinsic apoptosis),内因性アポトーシス(intrinsic apoptosis),オートファジー依存性細胞死(autophagy-dependent cell death),ネクロトーシス(necroptosis),フェロトーシス(ferroptosis),パイロトーシス(pyroptosis),リソソーム依存性細胞死(lysosome-dependent cell death),パータナトス(parthanatos)や免疫原性細胞死(immunogenic cell death)などがRCDに該当する.このRCDの詳細な制御機構の解明の進展とともに,内因性および外因性アポトーシス,オートファジー依存性細胞死,ネクロトーシス,フェロトーシスやパイロトーシスなどのRCDの制御機構へのセラミドの関与を示す報告が蓄積しつつある.
セラミドはセラミド合成酵素(ceramide synthase:CERS)により生成され,アシル鎖長が異なる多様なセラミド分子種(図1)が,6種の各CERSアイソザイム(CERS1–6)の基質特異性により生成される9, 10)(木原の稿を参照).セラミド分子種特異的な生物学的役割4)や特定のRCDへの特異的なセラミド分子種の関与も示唆されている11).本稿ではセラミド分子種に着目して,6種RCD(外因性アポトーシス,内因性アポトーシス,オートファジー依存性細胞死,ネクロトーシス,フェロトーシスおよびパイロトーシスを含む)の制御におけるセラミドの役割についてまとめた.
1972年Kerrらにより提唱されたアポトーシス12)は,カスパーゼ依存的に進行するRCDである.このアポトーシスには,外因性アポトーシスと内因性アポトーシスが存在する.外因性アポトーシスはdeath receptorsを介した細胞内シグナル伝達により,内因性アポトーシスはDNA損傷や代謝性ストレスから生じるミトコンドリア外膜透過性の亢進により誘導される.これらの細胞死を導くtumor necrosis factor(TNF)-α, Fasリガンド,抗がん剤,放射線および酸化ストレスなどへの細胞応答において,セラミドはセカンドメッセンジャーとして機能すると考えられている13, 14).
セラミド分子種に着目した研究から,C16:0-セラミドを含む長鎖セラミドはアポトーシス誘導性であることを示すエビデンスが蓄積しつつある.ここでは,セラミド分子種とアポトーシス制御に関する知見をまとめる(図2).
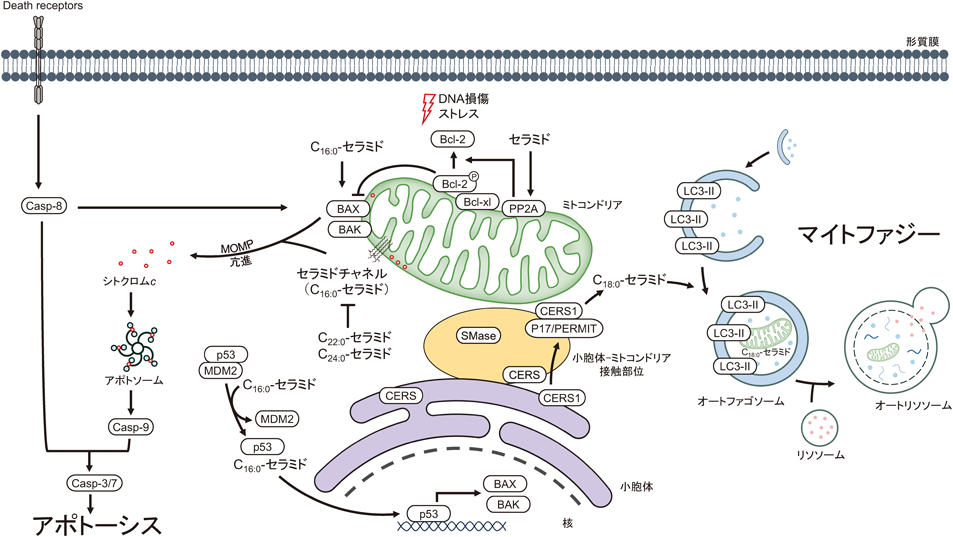
Casp:caspase, CERS:ceramide synthase, MAM:mitochondria-associated membrane, MDM2:mouse double minute protein 2, MOMP:mitochondrial outer membrane permeability, PP2A:protein phosphatase 2A, SMase:sphingomyelinase.
天然型セラミドは,多くの場合,炭素数16以上を含む脂肪酸が側鎖に結合しているため,水への溶解性や細胞膜透過性は低い.天然型セラミドの水溶性と細胞膜透過性を高めるため,側鎖の脂肪酸鎖を短鎖に変換した短鎖セラミドが人工合成された(図1).これらの短鎖C2:0-,C6:0-やC8:0-セラミドは細胞膜透過性を示すことから,アポトーシス研究において汎用され,細胞死制御の分子メカニズム解明に大きく寄与している1, 13, 15, 16).短鎖セラミドに応答した細胞死では,アポトーシスの生化学的特徴を示すDNAの断片化,カスパーゼの活性化,ミトコンドリアから細胞質ゾルへのシトクロムcの放出が観察される.これらの機序解明の中で,セラミドの標的細胞小器官や標的分子が同定されている13, 14).また,細胞内に取り込まれた短鎖セラミドはスフィンゴシン,グルコシルセラミド,セラミド1-リン酸やスフィンゴミエリンへと代謝されることから,この代謝を考慮した上でセラミド依存的な細胞死制御の機序解明も進んでいる17).細胞内に取り込まれた短鎖C6:0-セラミドは,セラミダーゼの加水分解を受け,スフィンゴシンへと変換される.さらにスフィンゴシンはCERSの触媒作用を介して,長鎖または極長鎖セラミドへと代謝される17).細胞内でのセラミド生成において,外因性セラミドまたは内因性セラミドからの新たなセラミド生成はそれぞれリサイクリング経路またはサルベージ経路として呼ばれている.これらの経路を経て生成されたセラミドの生物学的役割が明らかにされている17)(谷口の稿を参照).TakedaらはC6:0-セラミドのリサイクリング経路から生じた新たなセラミド分子種がアポトーシス誘導に寄与すること,一方でC2:0-セラミドはセラミダーゼにより加水分解されないため,リサイクリング経路を介した新たなセラミド生成が生じないことを示した18).これらは,短鎖セラミドを用いた研究を展開する上で重要な生化学的知見である.短鎖セラミドは,必ずしも細胞内で天然のセラミドと同じ代謝を受けるわけではないため,セラミドの生物学的応答においても両者に差異が生じる可能性がある.
アポトーシスの内因性経路において,ミトコンドリアからのシトクロムcの放出がアポトーシス誘導の引き金となる.シトクロムcはミトコンドリア外膜透過性の亢進により細胞質ゾルへと放出され,カスパーゼ-9や他の因子と複合体(アポトソーム)を形成する8, 19).このアポトソーム形成により活性化するカスパーゼ-9はアポトーシス実行分子カスパーゼ-3/7を活性化体へと変換する.これら一連の反応において,セラミドはシトクロムcの放出を促進し,その作用様式は多様である.
Bcl-2ファミリーには,ミトコンドリア外膜孔を形成するBAXやBAKなどのアポトーシス性分子,Bcl-2やBcl-xlなどの抗アポトーシス性分子があり,これらがミトコンドリア外膜透過性を制御することが知られている20, 21).BAXは分子内コンホメーション変化により活性化し,このコンホメーション変化はC16:0-セラミドによって誘導されること22),さらにセラミドと活性化型BAXはミトコンドリア外膜透過性を相乗的に高めること23)が報告されている.
Bcl-2は分子内70番目セリン残基のリン酸化により翻訳後調節されており,このリン酸化はBcl-2の抗アポトーシス作用を増強する24).この抗アポトーシス作用はセラミドにより減弱する.セリン/トレオニンプロテインホスファターゼであるprotein phosphatase 2A(PP2A)はceramide-activated protein phosphatasesとして同定され,短鎖セラミドはPP2Aのミトコンドリアへの移行とBcl-2の脱リン酸化に関わることが判明している25).
短鎖C2:0-セラミドを単離ミトコンドリアに添加すると,ミトコンドリアからシトクロムcが放出される26).この作用機序解明を目指した研究において,C2:0-セラミドまたはC16:0-セラミドはミトコンドリア膜でセラミドチャネルを形成することが示された27).このチャネルは,ミトコンドリア膜間腔から分子量約60,000を限度としたタンパク質を放出できる特徴を有しており,シトクロムc(分子量約12,000)の放出を担う可能性が示された28).また,これらの報告ではアポトーシス非誘導性ジヒドロセラミドのミトコンドリア外膜透過性への作用は微弱である.一方,C22:0-やC24:0-セラミドのような極長鎖セラミド分子種は無細胞系においてC16:0-セラミドのシトクロムc放出作用に対して拮抗することが判明している29).ミトコンドリアを起点とするアポトーシスの制御を理解する上で,セラミド分子種の構造特性に着目する必要があると思われる.
ミトコンドリア表面の5–20%が小胞体に近接(10–25 nm)しており,この小胞体–ミトコンドリア接触ゾーン(endoplasmic reticulum–mitochondria contact zones)には,CERSなどの脂質合成タンパク質や輸送タンパク質が局在する30–32).Mitochondria-associated membrane(MAM)とは小胞体–ミトコンドリア接触ゾーンを形成する小胞体内領域を生化学的に分離した膜画分を意味する33).2020年,Mignardらは,マウス肝臓から単離されたMAM画分中に含まれるスフィンゴ脂質量を質量分析法により解析した32).その結果,小胞体膜画分に匹敵するセラミドおよびスフィンゴミエリン量(pmol/mgタンパク質量)がMAM画分中に存在することが示された.ただし,この小胞体膜画分中にGM130(ゴルジ体マーカー)がわずかながら検出されることから,他の細胞小器官の混入にも留意する必要がある.また,単離ミトコンドリア画分中のC16:0-セラミド分子種量は他画分より多いことが判明した.さらにヒトがん細胞を用いた系において,アポトーシス誘導剤スタウロスポリン刺激下,細胞内C16:0-セラミド量の増加,ミトコンドリア画分中のセラミド量の増加やMAM画分中の酸性スフィンゴミエリナーゼ活性の上昇とスフィンゴミエリン量の減少が先行し,その後,ミトコンドリアからシトクロムcが放出された.これらの結果から,MAM画分中の酸性スフィンゴミエリナーゼの加水分解活性により生じたセラミド(おそらくC16:0-セラミド)がミトコンドリアに輸送され,ミトコンドリア外膜透過性の亢進に寄与したものと思われる.MAM画分では,リソソーム性タンパク質lysosome-associated membrane protein-2(LAMP2)やミトコンドリア外膜タンパク質Tom20が検出されたことから,酸性スフィンゴミエリナーゼ活性の検出は生化学的分画過程におけるリソソームのコンタミに由来する可能性も考えられる.しかしながら,スタウロスポリンによるアポトーシス誘導下,その活性は上昇することから,リソソームとMAMとの接触が亢進した可能性も考えられる.
放射線曝露への細胞応答において,セラミドがアポトーシス誘導を促進する34).線虫Caenorhabditis elegansの生殖細胞において,CERSが放射線によるアポトーシス誘導に必要であることが示された35).放射線曝露下HeLa細胞では,CERS5またはCERS6の過剰発現はアポトーシス誘導を促進し,逆にCERS2では抑制された36).これらより,CERS5およびCERS6から生成される長鎖セラミドはアポトーシス性,CERS2から生成される極長鎖セラミドは抗アポトーシス性である可能性が示された.また,この細胞応答において,ミトコンドリアでのC16:0-セラミド蓄積が観察されることから,ミトコンドリアを起点としたアポトーシス誘導へのC16:0-セラミドの関与が考えられる.また,オリゴデンドロサイトにおいても,グルタミン酸誘導性アポトーシスへのCERS6依存的なC16:0-セラミド生成の関与が示されている37).
がん抑制タンパク質p53は遺伝毒性などへの応答により活性化する転写因子であり,アポトーシス誘導性分子として知られている.細胞のストレス応答において,p53依存的なセラミド生成38)やセラミド依存的なp53活性化39)が報告されている.p53依存的なセラミド生成経路にCERS(CERS5およびCERS6)や中性スフィンゴミエリナーゼの関与が示されているが,その制御機構の詳細はいまだ不明である39, 40).
p53はE3ユビキチンリガーゼmouse double minute protein 2(MDM2)と複合体を形成しているため,p53のタンパク質量はプロテアソーム分解により低く抑えられている.細胞のストレス応答では,p53はその複合体から離脱し,安定化されることで,そのタンパク質量が増加する.Krupenkoらのグループは,C16:0-セラミドがp53のDNA結合部位に結合(Kd約60 nM)することで,p53-MDM2複合体からのp53の遊離とタンパク質としての安定性が亢進することを発見した41).またFatブロット法(疎水性メンブレンにスポットした脂質へのタンパク質の結合性を評価する生化学的手法)を用いた解析から,C16:0-セラミド以外のセラミド分子種(C12:0-,C14:0-,C18:0-,C20:0-,C22:0-,C24:0-およびC24:1-セラミド分子種)とp53との相互作用はなく,p53はC16:0-セラミド分子種に対して特異的に結合することが示された.これらの知見はセラミドによるp53制御の新たな機序を示すものであり,セラミド分子種特異的な構造生物学の解明につながる大きな前進である.
オートファジーは,栄養飢餓状態下,自己の細胞小器官やタンパク質を包み込んだオートファゴソームとリソソームが融合してオートリソソームを形成し,内容物を分解して再利用するプロセスである.オートファジーの進行は細胞生存または細胞死と関連する42).後者がオートファジー依存性細胞死(オートファジー関連細胞死)である.相反的であるが,セラミドはオートファジー依存的な細胞生存ならびにオートファジー依存性細胞死に関与することが知られている43).
短鎖セラミドを用いた研究から,セラミド依存的なオートファジー誘導機序が明らかにされている:i)AktおよびmTORシグナルの抑制44),ii)Bcl-2のリン酸化を介したBeclin1-Bcl-2複合体の解離45),iii)JNK/c-Jun経路の活性化によるBeclin1発現増加46),iv)アミノ酸トランスポーター(4F2he)やグルコーストランスポーター(GLUT1)の発現減少47).iii)およびiv)の報告では,セラミドのオートファジー依存性細胞死への関与が示された.
大麻に含まれる∆9-テトラヒドロカンナビノール(∆9-tetrahydrocannabinol)は,がん細胞においてセラミドde novo合成の刺激やオートファジー依存性細胞死の誘導を引き起こすことが知られている48–50).Velascoらの研究グループは,∆9-テトラヒドロカンナビノール刺激下において,小胞体でde novo合成されたジヒドロセラミドのオートファゴソーム/オートリソソームでの蓄積が細胞死誘導の要素の一つである可能性を示した51).これらをサポートする結果として,セラミドde novo合成阻害剤(ISP-1)によってリソソーム膜透過性の亢進(リソソーム性プロテアーゼの細胞質ゾルへの放出)および細胞死誘導が抑えられること,セラミド輸送タンパク質(ceramide transporter:CERT)52)を介した小胞体からゴルジ体へのセラミド輸送が抑えられること,およびオートファゴソーム濃縮画分でのジヒドロセラミド:セラミド比が上昇することが示された.これらの結果を考え合わせると,CERTやジヒドロセラミド不飽和化酵素の活性抑制がオートファジー依存性細胞死の誘導を促進する可能性が浮上する.また,小胞体でのジヒドロセラミドの蓄積が何らかにオメガソーム(オートファゴソームの前駆体)53)の形成を助長しているかもしれない.今後,さらなる研究の進展が期待される.
マイトファジーは,オートファジーによる選択的なミトコンドリアの分解である.C18:0-セラミド分子種が選択的にミトコンドリアのオートファゴソームとの融合を促進することで,マイトファジー誘導と細胞死を誘導する.その機序として,CERS1により生成されたC18:0-セラミドは,オートファゴソーム二重膜形成に必要なLC3-IIの疎水性ドメインに直接結合し,オートファゴソームの形成を誘導すること54),ならびに小胞体局在のCERS1はp17/PERMITの作用によりミトコンドリア外膜へ輸送され,そこでのC18:0-セラミドの生成がミトコンドリアのオートファゴソームとの融合に関わることが明らかにされた55)(図2).
ネクロトーシスはカスパーゼ非依存的に進行するプログラム化ネクローシスである8, 56–58).このプログラムの最終的な実行分子は,膜孔形成タンパク質mixed lineage kinase-domain like(MLKL)であり,当分子はreceptor-interacting protein kinase(RIPK)1ならびにRIPK3によりリン酸化され,分子内コンホメーションが変化し,多量体化する59, 60).この多量体(四量体または八量体)が形質膜へ移行し,その膜上で孔(イオンチャネル)として働くことで,細胞の膨潤や形質膜の破綻を来す60, 61).このようなネクロトーシス誘導経路において,マイクロドメイン62),極長鎖脂肪酸63, 64),イノシトールリン脂質65, 66),スフィンゴ脂質66, 67)の新たな機能が注目されている.ここでは,ネクロトーシス誘導下でのセラミド生成,MLKLの形質膜移行や多量体形成へのセラミドの関与について紹介する(図3).
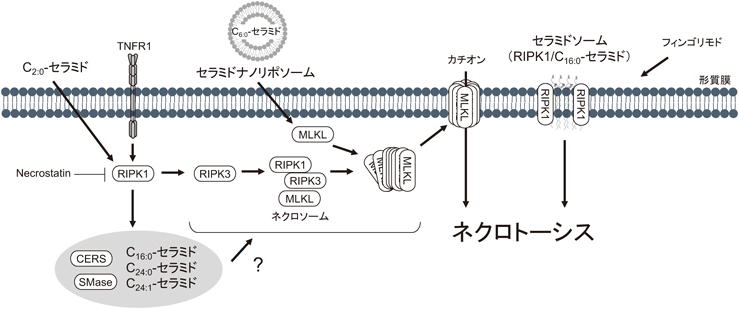
MLKL:mixed lineage kinase-domain like protein, RIPK:receptor-interacting protein kinase, TNFR1:tumor necrosis factor receptor 1.
Death receptorに分類されるTNFR1の刺激はカスパーゼ経路を活性化することで外因性アポトーシス誘導を促進するが,RIPKの活性化を抑えるカスパーゼ-8の阻害条件下ではネクロトーシス性細胞死が進行する58).このネクロトーシスのモデルは最も汎用されている.RIPK依存的なネクロトーシス経路では,まずネクロソーム(RIPK1/3およびMLKL複合体)が形成され,連続的にRIPKとMLKLが活性化する.TNFR1を介したアポトーシス誘導下ではスフィンゴミエリナーゼの活性化やセラミド生成が知られている.ところが,筆者らの知る限り,ネクロトーシス誘導下での酵素活性の変動を示す報告はなく,セラミド量の増加を示す研究が2報ある63, 68).これらの報告に共通してC16:0-,C24:0-およびC24:1-セラミド分子種が刺激応答において蓄積し,これらの蓄積はRIPK1阻害剤necrostatin処理により抑制されることが示された.これら共通の変化に加え,Sawaiら68)はRIPK1依存的なスフィンゴミエリン量の減少を観察しており,RIPK1がスフィンゴミエリナーゼの活性化に関与する可能性がある.Atilla-Gokcumenらのグループ63)は,複数のCERSアイソザイム(CERS1/3/5/6)の遺伝子発現が亢進することを示した.このことから,RIPK1依存的なセラミド蓄積へのCERSの関与も推察される.さらに同グループは脂肪酸伸長酵素ELOVLs(ELOVL1およびELOVL7)69, 70)の遺伝子発現上昇,極長鎖脂肪酸の蓄積,およびELOVL7のネクロトーシスへの関与を示した.ELOVL1はCERS2を介したC24:0-セラミドの産生に関連すること71)から,極長鎖セラミド分子種はネクロトーシスの誘導に関与する可能性が考えられる.今後,RIPK依存的なセラミド生成機序の解明やセラミド分子種特異的な生物学に関する研究の展開が望まれる.
短鎖セラミドを用いた細胞死の研究報告では,アポトーシスの解析が主である.ところが,2014年,頭頚部がん細胞株において,C2:0-セラミドによる細胞死はカスパーゼ非依存的であり,necrostatinにより抑えられることが報告された72).詳細な解析はないが,RIPK1依存性ネクロトーシス誘導へのセラミドの関与が初めて示された.
その後,C6:0-セラミドのリポソーム化製剤(セラミドナノリポソーム)は卵巣がん細胞においてアポトーシスではなく,ネクロトーシスを誘導することも示された73).生化学的特徴では,アポトーシスを示唆するカスパーゼ-3の活性化やPARPの分解は観察されず,ネクロトーシスを示唆するMLKLの多量体化と形質膜への移行が観察された.この細胞死誘導に対して,necrostatinやRIPK3 siRNAは影響を示さず,唯一MLKL siRNAが抑制作用を示したことから,セラミドナノリポソームはRIPK非依存的にMLKLを活性化すると考えられる.C6:0-セラミドの細胞内代謝や生化学的な機序は十分に解析されていないが,セラミドはMLKLの形質膜移行および膜孔形成に必要な多量体化に関与すると考えられ,今後,詳細な機序の解明が期待される.
スフィンゴイド塩基と構造が類似したフィンゴリモドは,RIPK3/MLKL非依存的,RIPK1依存的な細胞死を誘導することが報告されている74, 75).この細胞死においてセラミドソームと名づけられたRIPK1とC16:0-セラミドとの複合体が非筋細胞ミオシンII依存的に形質膜へと運ばれ,形質膜の破綻を来す孔として機能する可能性が提唱されている75).セラミドソームが孔形成を示す十分な証拠はないが,セラミドソームは形質膜上でセラミドが密に集積したプラットフォームの一つであると思われる.
パイロトーシス76, 77)はネクローシス様形態を示す細胞死であり,その誘導は細胞の炎症性応答に深く関連している.2015年にパイロトーシス実行の中心分子としてgasdermin(GSDM)ファミリー分子が発見されたこと78)から,パイロトーシス研究が飛躍的に進展している.生化学的な特徴として,このパイロトーシスでは,アポトーシスで観察されるカスパーゼの活性化やホスファチジルセリンの細胞表面への露出,GSDMのプロテオリシスによる活性化や形質膜孔の形成などがあげられる.GSDMファミリー分子の一つであるGSDMDが関与するパイロトーシスでは,炎症性応答により活性化されるカスパーゼ-1/4/5/11がGSDMDを切断し,その結果,N末端を保持する断片がオリゴマー化して形質膜へ移行する.その後,形質膜上で細孔を形成し,形質膜の破裂を引き起こす.このパイロトーシス制御機構へのセラミドの関与はほとんど明らかにされていないが,先駆的な二つの報告について概説する.
血管内皮細胞の障害の一つとしてNLRP3インフラマソーム依存的なパイロトーシスの関与が指摘されており,この細胞死へのセラミドの関与が報告された79).短鎖セラミド(C8:0-セラミド)は活性酸素種の蓄積に関わる分子やNLRP3の発現を高めることで,ヒト臍帯静脈内皮細胞においてパイロトーシスを誘導した.C8:0-セラミド代謝やセラミド標的分子について解析されていないが,パイロトーシス制御へのセラミドの関与を示唆するものである.
また,パイロトーシスはがんの新たな治療標的として注目されている.抗がん剤を用いた治療において,薬剤耐性化は深刻な課題である.この耐性化は,p53を介したカスパーゼ-3依存的なアポトーシスに抵抗性を示すことである.Suらは,抗がん剤耐性にβ5インテグリンが関わり,その経路がsrc-signal transducer and activator of transcription 3(STAT3)経路依存的に酸性セラミダーゼの発現を高めることで抗がん剤耐性の獲得につながることを示した80).おそらく,酸性セラミダーゼの加水分解活性がセラミドの蓄積を抑えることで抗がん剤耐性化に寄与していると考えられる.逆に酸性セラミダーゼの活性を抑えると,抗がん剤耐性細胞に対してパイロトーシスを誘導することが示された.酸性セラミダーゼはパイロトーシスを制御するスフィンゴ脂質代謝酵素の一つとして考えられる.
フェロトーシスは,非アポトーシス性の細胞死であり,鉄イオン依存的な活性酸素種の生成とリン脂質の過酸化により生じる細胞死として同定された81, 82).これまでに,このフェロトーシスの誘導に酸性スフィンゴミエリナーゼ83, 84),酸性セラミダーゼ85),セラミドキナーゼ86)やCERS687)の関与が指摘されている.しかしながら,報告数は限られている.フェロトーシスの誘導剤として知られているerastinはシスチン・グルタミン酸トランスポーター(system Xc)の阻害剤であり,この阻害作用により細胞内グルタチオンの量は低下し,リン脂質ヒドロペルオキシドグルタチオンペルオキシダーゼ4(GPX4)の抗酸化作用が減弱する.この結果,脂質過酸化物の蓄積によって細胞死が引き起こされる.このerastinへの細胞応答において,経時的なセラミド量の増加ならびに酸性スフィンゴミエリナーゼ活性の上昇が観察され,C16:0-ならびにC20:0-セラミド分子種選択的な増加が示された(C14:0-,C18:0-,C22:0-,C24:0-およびC24:1-セラミド量は変動しなかった)83).酸性スフィンゴミエリナーゼのノックダウンや薬理学的阻害によりセラミド生成ならびに細胞死は抑えられることから,酸性スフィンゴミエリナーゼのフェロトーシス誘導への関与が明らかになった.この機序として,NADPHオキシダーゼ依存的な活性酸素によって活性化した酸性スフィンゴミエリナーゼが何らかの作用によってGPX4のオートファジー性分解を促進したと考えられている.これらから,酸性スフィンゴミエリナーゼはフェロトーシスの制御分子の一つとして考えられ,酸性スフィンゴミエリナーゼにより生成されたC16:0-およびC20:0-セラミドはフェロトーシス誘導を担うセラミド分子種であるかもしれない.今後,フェロトーシス下でのセラミド代謝調節やセラミド分子種の生物学的役割を解き明かす必要がある.
各RCDにはそれぞれの仕組みがあり,仕組みを構成するタンパク質はプログラム的に特定の細胞小器官で機能することで細胞死が引き起こされる.RCDにおいて,セラミドは進行を制御する脂質性分子であり,その制御へのセラミド分子種選択的な関与がある.この制御には,個々のセラミド分子種の生成経路,輸送や生物学的特性が含まれるが,セラミド分子種の多様性とRCDの多様性が交わり,RCDにおけるセラミドの役割は想像以上に複雑である.この複雑さは,セラミド分子種の構造生物学的理解のさらなる進展によりひも解かれると期待される.
1) Obeid, L.M., Linardic, C.M., Karolak, L.A., & Hannun, Y.A. (1993) Programmed cell death induced by ceramide. Science, 259, 1769–1771.
2) Hannun, Y.A. (1996) Functions of ceramide in coordinating cellular responses to stress. Science, 274, 1855–1859.
3) Hannun, Y.A. & Obeid, L.M. (2008) Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol., 9, 139–150.
4) Hannun, Y.A. & Obeid, L.M. (2011) Many ceramides. J. Biol. Chem., 286, 27855–27862.
5) Yamaji, T. & Hanada, K. (2015) Sphingolipid metabolism and interorganellar transport: Localization of sphingolipid enzymes and lipid transfer proteins. Traffic, 16, 101–122.
6) Ogretmen, B. (2018) Sphingolipid metabolism in cancer signalling and therapy. Nat. Rev. Cancer, 18, 33–50.
7) Taniguchi, M. & Okazaki, T. (2021) Role of ceramide/sphingomyelin (SM) balance regulated through “SM cycle” in cancer. Cell. Signal., 87, 110119.
8) Galluzzi, L., Vitale, I., Aaronson, S.A., Abrams, J.M., Adam, D., Agostinis, P., Alnemri, E.S., Altucci, L., Amelio, I., Andrews, D.W., et al. (2018) Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ., 25, 486–541.
9) Mizutani, Y., Kihara, A., & Igarashi, Y. (2005) Mammalian Lass6 and its related family members regulate synthesis of specific ceramides. Biochem. J., 390, 263–271.
10) Levy, M. & Futerman, A.H. (2010) Mammalian ceramide synthases. IUBMB Life, 62, 347–356.
11) Alizadeh, J., da Silva Rosa, S.C., Weng, X., Jacobs, J., Lorzadeh, S., Ravandi, A., Vitorino, R., Pecic, S., Zivkovic, A., Stark, H., et al. (2023) Ceramides and ceramide synthases in cancer: Focus on apoptosis and autophagy. Eur. J. Cell Biol., 102, 151337.
12) Kerr, J.F., Wyllie, A.H., & Currie, A.R. (1972) Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer, 26, 239–257.
13) Ruvolo, P.P. (2003) Intracellular signal transduction pathways activated by ceramide and its metabolites. Pharmacol. Res., 47, 383–392.
14) Hannun, Y.A. & Obeid, L.M. (2018) Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol., 19, 175–191.
15) Bose, R., Verheij, M., Haimovitz-Friedman, A., Scotto, K., Fuks, Z., & Kolesnick, R. (1995) Ceramide synthase mediates daunorubicin-induced apoptosis: An alternative mechanism for generating death signals. Cell, 82, 405–414.
16) Hartfield, P.J., Mayne, G.C., & Murray, A.W. (1997) Ceramide induces apoptosis in PC12 cells. FEBS Lett., 401, 148–152.
17) Kitatani, K., Idkowiak-Baldys, J., & Hannun, Y.A. (2008) The sphingolipid salvage pathway in ceramide metabolism and signaling. Cell. Signal., 20, 1010–1018.
18) Takeda, S., Mitsutake, S., Tsuji, K., & Igarashi, Y. (2006) Apoptosis occurs via the ceramide recycling pathway in human HaCaT keratinocytes. J. Biochem., 139, 255–262.
19) Li, P., Nijhawan, D., Budihardjo, I., Srinivasula, S.M., Ahmad, M., Alnemri, E.S., & Wang, X. (1997) Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell, 91, 479–489.
20) Tsujimoto, Y. & Shimizu, S. (2000) VDAC regulation by the Bcl-2 family of proteins. Cell Death Differ., 7, 1174–1181.
21) Sharpe, J.C., Arnoult, D., & Youle, R.J. (2004) Control of mitochondrial permeability by Bcl-2 family members. Biochim. Biophys. Acta Mol. Cell Res., 1644, 107–113.
22) Kashkar, H., Wiegmann, K., Yazdanpanah, B., Haubert, D., & Kronke, M. (2005) Acid sphingomyelinase is indispensable for UV light-induced Bax conformational change at the mitochondrial membrane. J. Biol. Chem., 280, 20804–20813.
23) Ganesan, V., Perera, M.N., Colombini, D., Datskovskiy, D., Chadha, K., & Colombini, M. (2010) Ceramide and activated Bax act synergistically to permeabilize the mitochondrial outer membrane. Apoptosis, 15, 553–562.
24) Ruvolo, P.P., Deng, X., & May, W.S. (2001) Phosphorylation of Bcl2 and regulation of apoptosis. Leukemia, 15, 515–522.
25) Ruvolo, P.P., Deng, X., Ito, T., Carr, B.K., & May, W.S. (1999) Ceramide induces Bcl2 dephosphorylation via a mechanism involving mitochondrial PP2A. J. Biol. Chem., 274, 20296–20300.
26) Ghafourifar, P., Klein, S.D., Schucht, O., Schenk, U., Pruschy, M., Rocha, S., & Richter, C. (1999) Ceramide induces cytochrome c release from isolated mitochondria. Importance of mitochondrial redox state. J. Biol. Chem., 274, 6080–6084.
27) Siskind, L.J. & Colombini, M. (2000) The lipids C2- and C16-ceramide form large stable channels. Implications for apoptosis. J. Biol. Chem., 275, 38640–38644.
28) Siskind, L.J., Kolesnick, R.N., & Colombini, M. (2002) Ceramide channels increase the permeability of the mitochondrial outer membrane to small proteins. J. Biol. Chem., 277, 26796–26803.
29) Stiban, J. & Perera, M. (2015) Very long chain ceramides interfere with C16-ceramide-induced channel formation: A plausible mechanism for regulating the initiation of intrinsic apoptosis. Biochim. Biophys. Acta Biomembr., 1848, 561–567.
30) Bionda, C., Portoukalian, J., Schmitt, D., Rodriguez-Lafrasse, C., & Ardail, D. (2004) Subcellular compartmentalization of ceramide metabolism: MAM (mitochondria-associated membrane) and/or mitochondria? Biochem. J., 382, 527–533.
31) Grimm, S. (2012) The ER-mitochondria interface: The social network of cell death. Biochim. Biophys. Acta Mol. Cell Res., 1823, 327–334.
32) Mignard, V., Dubois, N., Lanoe, D., Joalland, M.P., Oliver, L., Pecqueur, C., Heymann, D., Paris, F., Vallette, F.M., & Lalier, L. (2020) Sphingolipid distribution at mitochondria-associated membranes (MAMs) upon induction of apoptosis. J. Lipid Res., 61, 1025–1037.
33) Vance, J.E. (1990) Phospholipid synthesis in a membrane fraction associated with mitochondria. J. Biol. Chem., 265, 7248–7256.
34) Haimovitz-Friedman, A., Kan, C.C., Ehleiter, D., Persaud, R.S., McLoughlin, M., Fuks, Z., & Kolesnick, R.N. (1994) Ionizing radiation acts on cellular membranes to generate ceramide and initiate apoptosis. J. Exp. Med., 180, 525–535.
35) Deng, X., Yin, X., Allan, R., Lu, D.D., Maurer, C.W., Haimovitz-Friedman, A., Fuks, Z., Shaham, S., & Kolesnick, R. (2008) Ceramide biogenesis is required for radiation-induced apoptosis in the germ line of C. elegans. Science, 322, 110–115.
36) Mesicek, J., Lee, H., Feldman, T., Jiang, X., Skobeleva, A., Berdyshev, E.V., Haimovitz-Friedman, A., Fuks, Z., & Kolesnick, R. (2010) Ceramide synthases 2, 5, and 6 confer distinct roles in radiation-induced apoptosis in HeLa cells. Cell. Signal., 22, 1300–1307.
37) Novgorodov, S.A., Chudakova, D.A., Wheeler, B.W., Bielawski, J., Kindy, M.S., Obeid, L.M., & Gudz, T.I. (2011) Developmentally regulated ceramide synthase 6 increases mitochondrial Ca2+ loading capacity and promotes apoptosis. J. Biol. Chem., 286, 4644–4658.
38) Dbaibo, G.S., Pushkareva, M.Y., Rachid, R.A., Alter, N., Smyth, M.J., Obeid, L.M., & Hannun, Y.A. (1998) p53-dependent ceramide response to genotoxic stress. J. Clin. Invest., 102, 329–339.
39) Fekry, B., Jeffries, K.A., Esmaeilniakooshkghazi, A., Ogretmen, B., Krupenko, S.A., & Krupenko, N.I. (2016) CerS6 is a novel transcriptional target of p53 protein activated by non-genotoxic stress. J. Biol. Chem., 291, 16586–16596.
40) Shamseddine, A.A., Clarke, C.J., Carroll, B., Airola, M.V., Mohammed, S., Rella, A., Obeid, L.M., & Hannun, Y.A. (2015) P53-dependent upregulation of neutral sphingomyelinase-2: Role in doxorubicin-induced growth arrest. Cell Death Dis., 6, e1947.
41) Fekry, B., Jeffries, K.A., Esmaeilniakooshkghazi, A., Szulc, Z.M., Knagge, K.J., Kirchner, D.R., Horita, D.A., Krupenko, S.A., & Krupenko, N.I. (2018) C(16)-ceramide is a natural regulatory ligand of p53 in cellular stress response. Nat. Commun., 9, 4149.
42) Denton, D., Nicolson, S., & Kumar, S. (2012) Cell death by autophagy: Facts and apparent artefacts. Cell Death Differ., 19, 87–95.
43) Dany, M. & Ogretmen, B. (2015) Ceramide induced mitophagy and tumor suppression. Biochim. Biophys. Acta Mol. Cell Res., 1853(10 Pt B), 2834–2845.
44) Schubert, K.M., Scheid, M.P., & Duronio, V. (2000) Ceramide inhibits protein kinase B/Akt by promoting dephosphorylation of serine 473. J. Biol. Chem., 275, 13330–13335.
45) Pattingre, S., Bauvy, C., Carpentier, S., Levade, T., Levine, B., & Codogno, P. (2009) Role of JNK1-dependent Bcl-2 phosphorylation in ceramide-induced macroautophagy. J. Biol. Chem., 284, 2719–2728.
46) Li, D.D., Wang, L.L., Deng, R., Tang, J., Shen, Y., Guo, J.F., Wang, Y., Xia, L.P., Feng, G.K., Liu, Q.Q., et al. (2009) The pivotal role of c-Jun NH2-terminal kinase-mediated Beclin 1 expression during anticancer agents-induced autophagy in cancer cells. Oncogene, 28, 886–898.
47) Guenther, G.G., Peralta, E.R., Rosales, K.R., Wong, S.Y., Siskind, L.J., & Edinger, A.L. (2008) Ceramide starves cells to death by downregulating nutrient transporter proteins. Proc. Natl. Acad. Sci. USA, 105, 17402–17407.
48) Galve-Roperh, I., Sanchez, C., Cortes, M.L., Gomez del Pulgar, T., Izquierdo, M., & Guzman, M. (2000) Anti-tumoral action of cannabinoids: Involvement of sustained ceramide accumulation and extracellular signal-regulated kinase activation. Nat. Med., 6, 313–319.
49) Salazar, M., Carracedo, A., Salanueva, I.J., Hernandez-Tiedra, S., Lorente, M., Egia, A., Vazquez, P., Blazquez, C., Torres, S., Garcia, S., et al. (2009) Cannabinoid action induces autophagy-mediated cell death through stimulation of ER stress in human glioma cells. J. Clin. Invest., 119, 1359–1372.
50) Velasco, G., Sanchez, C., & Guzman, M. (2012) Towards the use of cannabinoids as antitumour agents. Nat. Rev. Cancer, 12, 436–444.
51) Hernandez-Tiedra, S., Fabrias, G., Davila, D., Salanueva, I.J., Casas, J., Montes, L.R., Anton, Z., Garcia-Taboada, E., Salazar-Roa, M., Lorente, M., et al. (2016) Dihydroceramide accumulation mediates cytotoxic autophagy of cancer cells via autolysosome destabilization. Autophagy, 12, 2213–2229.
52) Hanada, K., Kumagai, K., Yasuda, S., Miura, Y., Kawano, M., Fukasawa, M., & Nishijima, M. (2003) Molecular machinery for non-vesicular trafficking of ceramide. Nature, 426, 803–809.
53) Hayashi-Nishino, M., Fujita, N., Noda, T., Yamaguchi, A., Yoshimori, T., & Yamamoto, A. (2010) Electron tomography reveals the endoplasmic reticulum as a membrane source for autophagosome formation. Autophagy, 6, 301–303.
54) Sentelle, R.D., Senkal, C.E., Jiang, W., Ponnusamy, S., Gencer, S., Selvam, S.P., Ramshesh, V.K., Peterson, Y.K., Lemasters, J.J., Szulc, Z.M., et al. (2012) Ceramide targets autophagosomes to mitochondria and induces lethal mitophagy. Nat. Chem. Biol., 8, 831–838.
55) Oleinik, N., Kim, J., Roth, B.M., Selvam, S.P., Gooz, M., Johnson, R.H., Lemasters, J.J., & Ogretmen, B. (2019) Mitochondrial protein import is regulated by p17/PERMIT to mediate lipid metabolism and cellular stress. Sci. Adv., 5, eaax1978.
56) Sun, L., Wang, H., Wang, Z., He, S., Chen, S., Liao, D., Wang, L., Yan, J., Liu, W., Lei, X., et al. (2012) Mixed lineage kinase domain-like protein mediates necrosis signaling downstream of RIP3 kinase. Cell, 148, 213–227.
57) Linkermann, A. & Green, D.R. (2014) Necroptosis. N. Engl. J. Med., 370, 455–465.
58) Tang, D., Kang, R., Berghe, T.V., Vandenabeele, P., & Kroemer, G. (2019) The molecular machinery of regulated cell death. Cell Res., 29, 347–364.
59) Wang, H., Sun, L., Su, L., Rizo, J., Liu, L., Wang, L.F., Wang, F.S., & Wang, X. (2014) Mixed lineage kinase domain-like protein MLKL causes necrotic membrane disruption upon phosphorylation by RIP3. Mol. Cell, 54, 133–146.
60) Meng, Y., Garnish, S.E., Davies, K.A., Black, K.A., Leis, A.P., Horne, C.R., Hildebrand, J.M., Hoblos, H., Fitzgibbon, C., Young, S.N., et al. (2023) Phosphorylation-dependent pseudokinase domain dimerization drives full-length MLKL oligomerization. Nat. Commun., 14, 6804.
61) Xia, B., Fang, S., Chen, X., Hu, H., Chen, P., Wang, H., & Gao, Z. (2016) MLKL forms cation channels. Cell Res., 26, 517–528.
62) Chen, X., Li, W., Ren, J., Huang, D., He, W.T., Song, Y., Yang, C., Li, W., Zheng, X., Chen, P., et al. (2014) Translocation of mixed lineage kinase domain-like protein to plasma membrane leads to necrotic cell death. Cell Res., 24, 105–121.
63) Parisi, L.R., Li, N., & Atilla-Gokcumen, G.E. (2017) Very long chain fatty acids are functionally involved in necroptosis. Cell Chem. Biol., 24, 1445–1454.e8.
64) Pradhan, A.J., Lu, D., Parisi, L.R., Shen, S., Berhane, I.A., Galster, S.L., Bynum, K., Monje-Galvan, V., Gokcumen, O., Chemler, S.R., et al. (2021) Protein acylation by saturated very long chain fatty acids and endocytosis are involved in necroptosis. Cell Chem. Biol., 28, 1298–1309.e7.
65) Dondelinger, Y., Declercq, W., Montessuit, S., Roelandt, R., Goncalves, A., Bruggeman, I., Hulpiau, P., Weber, K., Sehon, C.A., Marquis, R.W., et al. (2014) MLKL compromises plasma membrane integrity by binding to phosphatidylinositol phosphates. Cell Rep., 7, 971–981.
66) Zhang, X., Matsuda, M., Yaegashi, N., Nabe, T., & Kitatani, K. (2020) Regulation of necroptosis by phospholipids and sphingolipids. Cells, 9, 627.
67) Pilatova, M.B., Solarova, Z., Mezencev, R., & Solar, P. (2023) Ceramides and their roles in programmed cell death. Adv. Med. Sci., 68, 417–425.
68) Sawai, H., Ogiso, H., & Okazaki, T. (2015) Differential changes in sphingolipids between TNF-induced necroptosis and apoptosis in U937 cells and necroptosis-resistant sublines. Leuk. Res., 39, 964–970.
69) Kihara, A. (2012) Very long-chain fatty acids: Elongation, physiology and related disorders. J. Biochem., 152, 387–395.
70) Sassa, T. & Kihara, A. (2014) Metabolism of very long-chain Fatty acids: Genes and pathophysiology. Biomol. Ther. (Seoul), 22, 83–92.
71) Ohno, Y., Suto, S., Yamanaka, M., Mizutani, Y., Mitsutake, S., Igarashi, Y., Sassa, T., & Kihara, A. (2010) ELOVL1 production of C24 acyl-CoAs is linked to C24 sphingolipid synthesis. Proc. Natl. Acad. Sci. USA, 107, 18439–18444.
72) Zhu, W., Wang, X., Zhou, Y., & Wang, H. (2014) C2-ceramide induces cell death and protective autophagy in head and neck squamous cell carcinoma cells. Int. J. Mol. Sci., 15, 3336–3355.
73) Zhang, X., Kitatani, K., Toyoshima, M., Ishibashi, M., Usui, T., Minato, J., Egiz, M., Shigeta, S., Fox, T., Deering, T., et al. (2018) Ceramide nanoliposomes as a MLKL-dependent, necroptosis-inducing, chemotherapeutic reagent in ovarian cancer. Mol. Cancer Ther., 17, 50–59.
74) Saddoughi, S.A., Gencer, S., Peterson, Y.K., Ward, K.E., Mukhopadhyay, A., Oaks, J., Bielawski, J., Szulc, Z.M., Thomas, R.J., Selvam, S.P., et al. (2013) Sphingosine analogue drug FTY720 targets I2PP2A/SET and mediates lung tumour suppression via activation of PP2A-RIPK1-dependent necroptosis. EMBO Mol. Med., 5, 105–121.
75) Nganga, R., Oleinik, N., Kim, J., Selvam, S.P., De Palma, R., Johnson, K.A., Parikh, R.Y., Gangaraju, V., Peterson, Y., Dany, M., et al. (2019) Receptor-interacting Ser/Thr kinase 1 (RIPK1) and myosin IIA-dependent ceramidosomes form membrane pores that mediate blebbing and necroptosis. J. Biol. Chem., 294, 502–519.
76) Yu, P., Zhang, X., Liu, N., Tang, L., Peng, C., & Chen, X. (2021) Pyroptosis: Mechanisms and diseases. Signal Transduct. Target. Ther., 6, 128.
77) Lu, L., Zhang, Y., Tan, X., Merkher, Y., Leonov, S., Zhu, L., Deng, Y., Zhang, H., Zhu, D., Tan, Y., et al. (2022) Emerging mechanisms of pyroptosis and its therapeutic strategy in cancer. Cell Death Discov., 8, 338.
78) Shi, J., Zhao, Y., Wang, K., Shi, X., Wang, Y., Huang, H., Zhuang, Y., Cai, T., Wang, F., & Shao, F. (2015) Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature, 526, 660–665.
79) Liu, F., Zhang, Y., Shi, Y., Xiong, K., Wang, F., & Yang, J. (2022) Ceramide induces pyroptosis through TXNIP/NLRP3/GSDMD pathway in HUVECs. BMC Mol. Cell Biol., 23, 54.
80) Su, L., Chen, Y., Huang, C., Wu, S., Wang, X., Zhao, X., Xu, Q., Sun, R., Kong, X., Jiang, X., et al. (2023) Targeting Src reactivates pyroptosis to reverse chemoresistance in lung and pancreatic cancer models. Sci. Transl. Med., 15, eabl7895.
81) Dixon, S.J., Lemberg, K.M., Lamprecht, M.R., Skouta, R., Zaitsev, E.M., Gleason, C.E., Patel, D.N., Bauer, A.J., Cantley, A.M., Yang, W.S., et al. (2012) Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell, 149, 1060–1072.
82) Imai, H., Matsuoka, M., Kumagai, T., Sakamoto, T., & Koumura, T. (2017) Lipid peroxidation-dependent cell death regulated by GPx4 and ferroptosis. Curr. Top. Microbiol. Immunol., 403, 143–170.
83) Thayyullathil, F., Cheratta, A.R., Alakkal, A., Subburayan, K., Pallichankandy, S., Hannun, Y.A., & Galadari, S. (2021) Acid sphingomyelinase-dependent autophagic degradation of GPX4 is critical for the execution of ferroptosis. Cell Death Dis., 12, 26.
84) Du, Y.X., Zhao, Y.T., Sun, Y.X., & Xu, A.H. (2023) Acid sphingomyelinase mediates ferroptosis induced by high glucose via autophagic degradation of GPX4 in type 2 diabetic osteoporosis. Mol. Med., 29, 125.
85) Ung, J., Tan, S.F., Fox, T.E., Shaw, J.J., Taori, M., Horton, B.J., Golla, U., Sharma, A., Szulc, Z.M., Wang, H.G., et al. (2023) Acid ceramidase inhibitor LCL-805 antagonizes akt signaling and promotes iron-dependent cell death in acute myeloid leukemia. Cancers (Basel), 15, 5866.
86) Vu, N.T., Kim, M., Stephenson, D.J., MacKnight, H.P., & Chalfant, C.E. (2022) Ceramide kinase inhibition drives ferroptosis and sensitivity to cisplatin in mutant KRAS lung cancer by dysregulating VDAC-mediated mitochondria function. Mol. Cancer Res., 20, 1429–1442.
87) Li, D., Tian, L., Nan, P., Zhang, J., Zheng, Y., Jia, X., Gong, Y., & Wu, Z. (2023) CerS6 triggered by high glucose activating the TLR4/IKKbeta pathway regulates ferroptosis of LO2 cells through mitochondrial oxidative stress. Mol. Cell. Endocrinol., 572, 111969.

摂南大学大学院薬学研究科医療薬学専攻博士課程2年生.学士(薬学および文学).
1980年大阪府に生る.99年梅花女子大学卒業,2023年摂南大学薬学部卒業,同年4月大学院入学.
研究テーマと抱負セラミドによるネクロトーシス誘導の機序解明
ウェブサイトhttps://www.setsunan.ac.jp/~p-official/index.html
趣味ゴルフ.

摂南大学薬学部 教授.博士(薬学).
1974年和歌山市に生る.97年京都薬科大学卒業,2002年同大学院博士課程修了,同年サウスカロライナ医科大学博士研究員,07年同大学インストラクター,08年鳥取大学助教,12年東北大学助教,17年同大学講師,18年摂南大学講師,21年同大学准教授,24年同大学教授.
研究テーマと抱負(1) スフィンゴ脂質の生化学と創薬応用.
(2) ネクロトーシスにおけるセラミドの役割と創薬応用.
(3) 新たながん治療標的分子の探索.
ウェブサイトhttps://www.setsunan.ac.jp/~p-official/index.html
趣味魚釣り.

This page was created on 2024-06-28T17:21:31.861+09:00
This page was last modified on 2024-08-19T08:50:42.000+09:00
このサイトは(株)国際文献社によって運用されています。