原核生物においては,スフィンゴ脂質(セラミド関連脂質)の生合成は,一部の細菌では知られているものの(生城らの稿を参照),すべての真核生物はセラミドを合成し,親水性頭部を持った複合スフィンゴ脂質を生体膜脂質の一構成成分として持つ.真核微生物の中でも出芽酵母は,スフィンゴ脂質の生合成酵素遺伝子の同定とスフィンゴ脂質の代謝制御機構の解明において多大な貢献をしてきた.たとえば,すべての真核生物で共通して存在するスフィンゴイド塩基からセラミドの生合成に至る経路で機能するセリンパルミトイル転移酵素(SPT),3-ケトジヒドロスフィンゴシン還元酵素,セラミド合成酵素の遺伝子クローニングは,出芽酵母が端緒となっている1–3).また,Orm1, Orm2を介したSPTの活性制御機構も出芽酵母で最初に発見されており,哺乳動物におけるORMDLタンパク質を介したスフィンゴ脂質生合成制御機構の解明にもつながっている4).それだけでなく,スフィンゴ脂質の分解代謝系(セラミドの分解,リン酸化を介したスフィンゴイド塩基のアシルCoAへの代謝経路等)に関わる酵素遺伝子の発見に関しても出芽酵母の研究が多大な貢献をしてきている5–8).一方で,出芽酵母の複合スフィンゴ脂質の構造バリエーションは哺乳動物よりも非常にシンプルである.このことを利用して,複合スフィンゴ脂質の構造多様性の生理的意義を理解するための研究も多く行われてきている.本稿では,主に出芽酵母の分子遺伝学を基盤とした複合スフィンゴ脂質の微細構造と生理機能の連関性の研究に関して概説する.
2. 真菌におけるセラミド,複合スフィンゴ脂質の構造の特徴
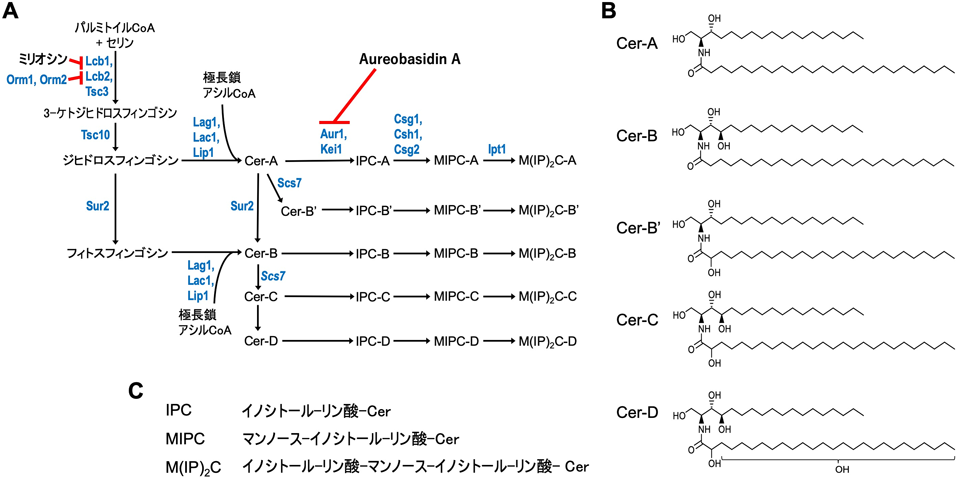
真菌では,C-4位に水酸基を持つフィトスフィンゴシンと呼ばれるスフィンゴイド塩基が主要なセラミドの基本骨格となる.また,哺乳動物と同じくC-4, 5位にトランス二重結合を持つスフィンゴシンも存在し,さらにC-8, 9位にトランス二重結合およびメチル基が付加した特徴的な構造もある(これらのスフィンゴイド塩基の構造は,Cryptococcus neoformans, Pichia pastoris, Candida albicans, Aspergillus oryzae等で確認されている9–12).本稿で主にとりあげる出芽酵母には,スフィンゴシンやそのメチル化された構造は存在しない).また,セラミドの脂肪酸部分のα位が水酸化されている場合が多い.複合スフィンゴ脂質としては,イノシトールリン酸を親水性頭部として持つイノシトールホスホセラミド(inositolphosphoceramide:IPC)が存在するが,スフィンゴミエリンは存在しない.出芽酵母Saccharomyces cerevisiaeにおいては,セラミドグルコシルトランスフェラーゼ(GlcT)遺伝子(ヒトにおけるUDP-glucose:ceramide glucosyltransferaseをコードするUGCGの真菌オルソログ)がないが,C. neoformans, P. pastoris, C. albicans等ではGlcT遺伝子が存在することでグルコシルセラミドの合成が行われる9–11, 13).日本酒の製造においては,並行複発酵と呼ばれる形式で,麹菌と清酒酵母を共存させデンプンの糖化とアルコール発酵を同時進行で行う.このとき,麹菌から放出されるグルコシルセラミドを清酒酵母が取り込むことで,アルカリ耐性を獲得するという報告がある14).実際,グルコシルセラミド,GlcT遺伝子を持つ酵母と持たない酵母,合計90種のアルカリ条件(pH 7.0–10.5)における生育能力を比較したところ,グルコシルセラミド,GlcT遺伝子の存在の有無とアルカリ耐性に相関があることが示されている15).また,C. neoformansではグルコシルセラミドが欠損すると宿主での感染能が低下することが報告されており,この原因も宿主の環境内におけるpH適応が関係しているとされている16).これらのことから,グルコシルセラミドの存在が酵母のpH適応に重要であることが示唆されている.出芽酵母(以下,単に出芽酵母と記した場合はS. cerevisiaeを指す)では,IPCにマンノースが付加してマンノシルイノシトールホスホセラミド(mannosyl-inositolphosphoceramide:MIPC),さらにMIPCにイノシトールリン酸が再度付加することでマンノシルジイノシトールホスホセラミド(mannosyl-diinositolphosphoceramide:M(IP)2C)が生合成される.一方で,分裂酵母Schizosaccharomyces pombeでは,MIPCまでしか生合成されない17).分裂酵母に出芽酵母のM(IP)2C生合成酵素遺伝子(IPT1)を発現させると,M(IP)2Cが細胞内で生合成されるようになるが,顕著な細胞死が観察される.このことは,種特異的な複合スフィンゴ脂質を別の種で発現させると,毒性を発揮してしまう場合があることを示している18).このように真菌間での異なる複合スフィンゴ脂質の組成が,生育や環境適応と密接に関係することが示唆されている.
3. 出芽酵母のセラミド,複合スフィンゴ脂質の生合成経路
出芽酵母におけるスフィンゴ脂質生合成の最初のステップは,哺乳動物と同じくセリンとアシルCoAの縮合反応であり,この反応を触媒するセリンパルミトイル転移酵素をコードする遺伝子(LCB1, LCB2)は,スフィンゴイド塩基要求性変異株の解析から初めて同定された1).その後,3-ケトジヒドロスフィンゴシン還元酵素(Tsc10),セラミド合成酵素(Lag1, Lac1)の作用を経てジヒドロセラミド(セラミド-A)が生合成される(図1A).Lag1, Lac1は極長鎖アシルCoA(極長鎖:≧C21)を基質として好むため,出芽酵母のセラミドの脂肪酸鎖長はほとんどがC26となる.これはC12からC24を中心として脂肪酸鎖長にバリエーションがある哺乳動物のセラミドとは大きく異なる構造的特性である.出芽酵母のセラミドは,スフィンゴイド塩基と脂肪酸部分の水酸化の違いによって,A, B, B′,C, Dの5種類に分類される(図1B).セラミド-Aはジヒドロスフィンゴシンと(非水酸化)脂肪酸から構成される.脂肪酸部分のほとんどはC26:0の飽和極長鎖脂肪酸である.セラミド-Aから,スフィンゴ脂質C4-ヒドロキシラーゼ(Sur2)によってジヒドロスフィンゴシン部分のC-4位が水酸化されセラミド-Bが,スフィンゴ脂質α-ヒドロキシラーゼ(Scs7)によって脂肪酸部分のC-2位が水酸化されセラミド-B′がそれぞれ生成される.セラミド-Cは,Sur2とScs7の両方の水酸化を受けている(図1).出芽酵母ではCタイプが最も多い19).なお,Sur2がジヒドロスフィンゴシンをフィトスフィンゴシンに変換し,これがセラミド-Bになる経路も存在すると思われる.セラミド-Dでは,セラミド-Cの脂肪酸部分の不明な位置が水酸化を受けている(図1).セラミド-Cからセラミド-Dへの変換を触媒するヒドロキシラーゼの分子実体は不明であるが,細胞内Cu2+トランスポーターをコードするCCC2の欠損でセラミド-Dが消失することから,その生合成にはCu2+が必要であることが示唆されている20).セラミドまでの生合成は小胞体で行われ,以降の複合スフィンゴ脂質への変換は,ゴルジ体で起こる.セラミドの小胞体からゴルジ体への輸送には,小胞体依存的および非依存的な経路が存在していると考えられており,後者の経路に関与するセラミド輸送タンパク質については複数の候補遺伝子がある21–24).ゴルジ体に輸送されたセラミドは,IPC合成酵素Aur1によってIPCに変換され,Csg1, Csh1によってMIPC, Ipt1によってM(IP)2Cに順次変換されていく(図1A, C)19, 25–27).Kei1はAur1, Csg2はCsg1, Csh1の調節サブユニットとしてそれぞれ機能する26, 28).以上のような合成経路を介して,複合スフィンゴ脂質の構造多様性が構成され,出芽酵母では15種類のサブタイプが生合成されうる(図1).
4. 特定の複合スフィンゴ脂質サブタイプとホスファチジルセリンの機能的相互作用
AUR1遺伝子にコードされるIPC合成酵素の阻害は,すべての複合スフィンゴ脂質の減少とセラミドの異常蓄積をもたらすことで,重篤な生育阻害を引き起こす.我々はIPC合成酵素阻害剤aureobasidin A(図1A)に対する高感受性を引き起こす変異株を出芽酵母遺伝子ノックアウトライブラリーからスクリーニングし,ホスホイノチシド脱リン酸化酵素をコードするSAC1の欠損株を同定した29).SAC1欠損は,複合スフィンゴ脂質の構造多様性を規定する遺伝子であるCSG1, CSG2, SCS7,もしくはIPT1との二重欠損によって合成致死(二つ以上の遺伝子の変異の組合わせで致死となる現象)となることがわかり,特定の複合スフィンゴ脂質とSac1との機能的相互作用が示唆された.ERで生合成されたホスファチジルセリン(PS)は,オキシステロール結合タンパク質Osh6/7を介して形質膜へと輸送されるが,その際ホスファチジルイノシトール4-リン酸のOsh6/7による形質膜からERへの交換輸送が必要となる.Sac1は,ERでホスファチジルイノシトール4-リン酸を分解することでPSの輸送系の維持において重要な役割を果たしており,SAC1欠損株ではPS量が減少することが報告されている30–32).SAC1と複合スフィンゴ脂質代謝酵素遺伝子の二重変異による合成致死は,SAC1欠損によるPS減少が原因の一つになることがわかった.また,PS合成酵素(遺伝子名CHO1)と複合スフィンゴ脂質代謝酵素の間でも遺伝学相互作用があり,二つの異なる膜脂質の代謝異常による合成致死を見いだした29).CHO1を発現抑制したCSG2欠損株では,初期エンドソームからゴルジ体への小胞輸送系に損傷が出ることがわかり,PSと特定の複合スフィンゴ脂質が協調的に特定の小胞輸送を制御することがわかった33).
5. MIPC生合成酵素の欠損と遺伝学的相互作用を示す遺伝子の探索
IPCからMIPCへの生合成の欠損は,MIPCとM(IP)2Cの完全な消失だけでなく,IPCの異常蓄積を引き起こす.CSG2の変異株は,100 mMのCaCl2に対して高感受性を示す株として同定された34).この高感受性の原因としては,CSG2欠損株の形質膜もしくは細胞壁に対して,Ca2+が不可逆的な損傷をもたらすことで生じることが考えられているが,詳細は不明である20).このCaCl2高感受性を抑制できるサプレッサー変異として,スフィンゴ脂質ヒドロキシラーゼ遺伝子(SUR2, SCS7),SPT遺伝子(LCB1, LCB2),3-ケトスフィンガニン還元酵素遺伝子(TSC10),エノイル還元酵素遺伝子(TSC13;極長鎖脂肪酸の生合成に関与)が同定されている35).これらの変異は,スフィンゴ脂質の総量の低下,もしくはIPC-Cの消失につながるものであり,CSG2変異によるCa2+高感受性は,MIPC生合成の低下によるIPC-Cの異常蓄積に起因すると考えられている.すなわち,IPC-Cの異常蓄積下では,形質膜もしくは細胞壁がCa2+による影響を受けやすくなることが想定される.MIPC生合成欠損株では窒素源飢餓下での迅速な細胞死がみられるが,これもIPC-C異常蓄積が原因であることが示唆されている36, 37).我々は,MIPC生合成の新たな生理的意義を見いだす目的で,CSG1, CSH1欠損との多重欠損によって合成致死を示す遺伝子を,出芽酵母遺伝子ノックアウトライブラリーを用いたsynthetic genetic array解析38)で探索し16個の遺伝子を見いだした39).この中には細胞壁の糖タンパク質糖鎖であるマンナンの合成に関与する糖転移酵素遺伝子が複数含まれていたことから,細胞壁とMIPC生合成との機能的連関性が示唆された.MIPC生合成欠失とマンナン生合成不全の二重代謝異常による合成生育損傷は,IPC-Cの蓄積を止める変異でも抑制はできなかった.また,MIPCからM(IP)2Cへの変換の欠失とマンナン生合成不全では生育損傷はみられなかった.すなわち,この遺伝学的相互作用はIPCの蓄積やM(IP)2Cの消失で起こるのではなく,MIPCの消失によって引き起こされることが示唆された.MIPC,マンナン二重代謝異常株は,細胞壁溶解酵素であるザイモリエースに対する感受性が顕著に増大しており,細胞壁の機能損傷が起きていることが示唆された39).また,MIPC生合成酵素欠損とエルゴステロール生合成経路の中間段階で働くスクアレン合成酵素の発現抑制でも細胞壁機能異常が顕著に促進されることも見いだした40).MIPC,エルゴステロールの二重代謝異常株では細胞壁の異常感知センサーであるWsc1, Wsc2を介して細胞壁修復シグナル[CWI(cell wall integrity)経路]が恒常的に活性化されており,酵母はこの変異株において常に細胞壁の機能補填をしようとしていることもわかった.これらの結果は,MIPCとエルゴステロールが協調的に細胞壁の機能維持に関与していることを示唆している40).MIPCの消失でなぜ細胞壁に機能損傷が生じるのかは不明であるが,細胞壁の必須成分であるGPIアンカー型タンパク質の細胞内輸送異常が起きている可能性が考えられる.またごく最近,分裂酵母においてMIPCの異常蓄積が細胞壁構成成分のβ-グルカンの異常分布を引き起こすことも報告されており,細胞内で適正なMIPC量を保つことが,正常な細胞壁機能の維持に重要であることも示唆されている41).
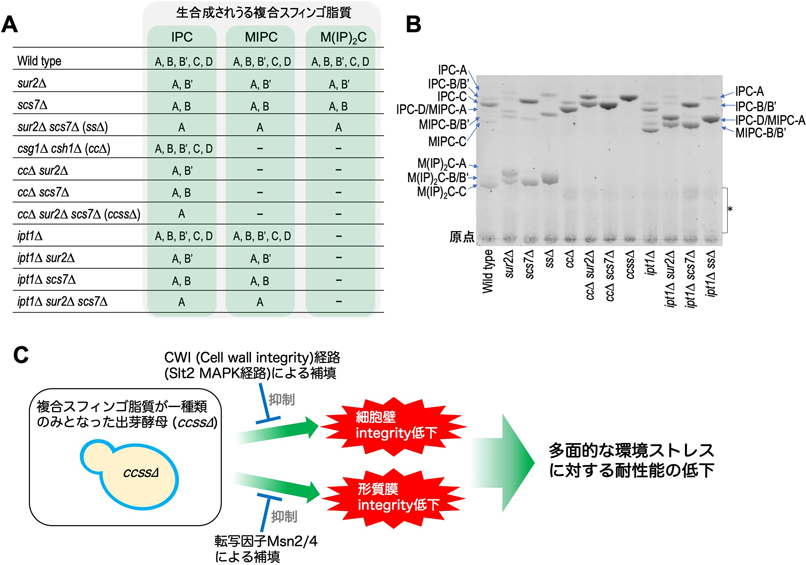
6. 複合スフィンゴ脂質構造多様性破綻による多面的環境ストレス高感受性
複合スフィンゴ脂質の構造多様性を規定する酵素遺伝子(CSG1, CSH1, SUR2, SCS7, IPT1)の欠損株における表現型はいくつか知られていたが,断片的な情報にとどまっていた.我々は出芽酵母における複合スフィンゴ脂質の構造多様性の生理学的意義を総合的に調べる目的で,複合スフィンゴ脂質構造多様性破綻ライブラリー(SUR2, SCS7, CSG1, CSH1, IPT1の欠損を組み合わせることで,さまざまな複合スフィンゴ脂質サブタイプが抜け落ちた計11種類の変異株ライブラリー)を作製し(図2A, B),それらの環境ストレス耐性能の評価を行った42).その結果,複合スフィンゴ脂質の構造多様性が限定されればされるほど,温度,浸透圧,有機酸,アルカリpH,種々の薬剤等の多様なストレスに対する感受性が増大する傾向が観察された.すなわち,複合スフィンゴ脂質が1種類のみとなった変異株[csg1∆ csh1∆ sur2∆ scs7∆(ccss∆株)]は,多くの環境ストレスに対して最も強い感受性を示すことがわかった.ccss∆株のストレス高感受性は,出芽酵母の典型的なストレス応答経路[HOG経路(後述),CWI経路,Rim101経路]の損傷で起きているものではないことが示唆された.さらにccss∆株では,CWI経路のMAPキナーゼであるSlt2やさまざまなストレス耐性に関わるジェネラルストレス応答転写因子であるMsn2/4が,ストレスをかけていない通常の培養条件における生育維持に必要となり,ccss∆株によって引き起こされるストレス耐性能低下に対しても補填をしていることがわかった.また,SLT2欠損はccss∆株のザイモリエース感受性を促進し,MSN2/4欠損は形質膜の透過性増大と流動性変化をもたらすことが判明し,Slt2およびMsn2/4がccss∆株の細胞壁および形質膜インテグリティの維持にそれぞれ寄与していることも示唆された.以上のことより,複合スフィンゴ脂質の構造多様性の限定は,細胞壁,形質膜といった細胞表面の環境の異常を介して多面的なストレス高感受性をもたらすことが考えられた.さらに出芽酵母はCWI経路やMsn2/4を介した転写応答によって,これらの異常に対する機能補填をしていることが示唆された(図2C)42).
7. 異常環境下におけるスフィンゴ脂質代謝変動とその生理的意義
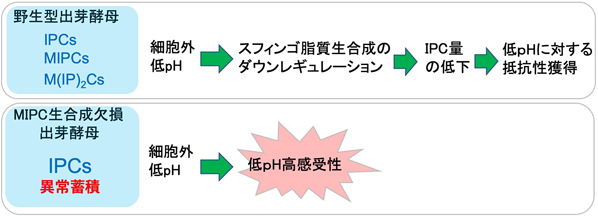
出芽酵母は基本的に弱酸性(pH 5.5前後)の環境を好み,そこからpHが大きく外れることで生育に重篤な影響が生じる.我々は,MIPC生合成欠損株(csg1∆ csh1∆株)が低pH(pH 2.5–3.5)条件下で,野生株と比較して生育能,生存率が大きく低下することを見いだした43).この表現型は,スフィンゴ脂質の総量の低下やSUR2, SCS7欠損によって回復することから,MIPC生合成の消失によるIPC-Cの異常蓄積によるものであることが示唆された.csg1∆ csh1∆株の低pH高感受性を抑制するサプレッサー変異を,トランスポゾン挿入酵母染色体ライブラリーを用いて探索した結果,形質膜のエルゴステロールの小胞体への輸送に関与するLAM1, SIP3, YSP2遺伝子44)を同定した.csg1∆ csh1∆株にLAM1, SIP3, YSP2をすべて欠損させると細胞表面のエルゴステロールの量の増大が確認され,MIPC生合成欠失による低pH高感受性は形質膜のエルゴステロールで補填できることが示された.また,エルゴステロール生合成の最終段階に関わるERG2, ERG6の欠損がcsg1∆ csh1∆による低pH高感受性を促進することもわかった43).
出芽酵母は,低pH下で自発的にIPCの代謝を制御している可能性はないだろうか? このことを調べるために,pH 2.5に野生型株をさらしたときの複合スフィンゴ脂質の組成パターンを解析した.その結果,低pH条件に3時間さらした後では,IPC量が半分以下に減少することが確認された.また,Lcb1, Aur1の発現量増大,Orm2の発現量低下もそれぞれ観察された.AUR1過剰発現およびORM1/ORM2を欠失した変異株では,低pH下での生育能が大きく低下していた.これらのことから,酵母は細胞外が低pHになると自発的にIPC量を低下させることで,環境適応をしていることが示唆された(図3)43).
液胞H+-ATPase(V-ATPase)は,サイトゾルから液胞にH+を取り込むことで,細胞内pHの恒常性を維持している45).我々は,V-ATPase欠損株が複合スフィンゴ脂質の生合成抑制に対して高感受性を示すことを見いだした46).また,V-ATPase欠損株では,IPC量低下,MIPC, M(IP)2C量増大,セラミド部分の水酸化の低下という複合スフィンゴ脂質のダイナミックな組成パターン変動が観察された.これらの変化を抑制する変異をV-ATPase欠損株に導入したところ,pH 7.2(V-ATPase欠損株の液胞におけるpHの低下を促進)の条件下での生育が顕著に低下した.これらのことより,細胞内のpHの制御が破綻すると,出芽酵母は自身の複合スフィンゴ脂質の組成を改変することで生き延びようとしていることが示唆された.
エンドサイトーシス,アクチン細胞骨格の制御に関わるamphiphysinファミリータンパク質遺伝子,RVS167, RVS161の欠損株でみられる典型的表現型である高温およびNaCl高感受性は,複合スフィンゴ脂質の代謝酵素遺伝子の変異で回復することが知られている47).我々は,RVS167欠損株ではOrm2の発現量が増大することでスフィンゴ脂質のde novo生合成が低下していることを見いだした48).RVS167欠損株で,スフィンゴ脂質生合成をさらに抑制させると,RVS167欠損によるエンドサイトーシスの遅延やアクチン細胞骨格の異常が緩和され,逆にスフィンゴ脂質生合成を促進させると表現型が増強されることも確認された.これらのことから,出芽酵母はamphiphysinの異常下でスフィンゴ脂質の生合成を調節することで,異常状況に適応をしようとしていることが示唆された48).
このような一連の結果は,出芽酵母はさまざまな異常環境下でスフィンゴ脂質代謝を調節し,積極的にその組成パターンを変化させることで,環境適応をしていることを示している.
8. セラミド,複合スフィンゴ脂質の生合成の破綻に対する救済機構
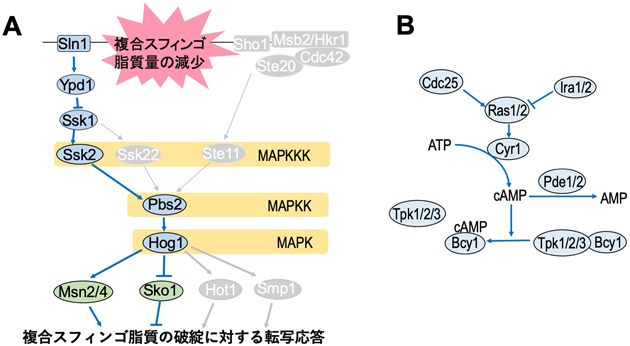
細胞内のスフィンゴ脂質,複合スフィンゴ脂質の総量の低下は,強力な生育阻害を誘導し,重篤になると細胞死に至る.我々は,AUR1発現抑制株における生育損傷を抑制できるサプレッサー変異を,トランスポゾン挿入出芽酵母染色体ライブラリーを用いて探索した結果,7個の遺伝子(SRB8, SSN3, RFX1, HTA1, XRN1, DCK1, RAS2)を同定した49).SSN3とSRB8はRNAポリメラーゼIIのメディエーター複合体をコードしており,Msn2を含むいくつかの転写因子の制御にも関与する50).MSN2は,AUR1発現抑制株の生育損傷を抑制するマルチコピーサプレッサー遺伝子としても同定されている49).DCK1は低分子量GTPaseのRho5のグアニンヌクレオチド交換因子をコードしており,Rho5は出芽酵母の高浸透圧耐性獲得と関連することが示唆されている51, 52).さらに先行研究で,複合スフィンゴ脂質の総量低下によって,高浸透圧応答(high osmolarity glycerol:HOG)経路と呼ばれるMAPキナーゼ経路が活性化されることが報告されている53).Msn2とMsn4はHOG経路下流の転写因子として働く54).これらのことを総合すると,HOG経路と複合スフィンゴ脂質の密接な連関性が考えられた.HOG経路のMAPキナーゼをコードするHOG1を欠損させると,AUR1もしくはLCB1発現抑制株における生育損傷が促進された49).一方で,HOG経路のMAPキナーゼキナーゼ遺伝子であるPBS2の過剰発現で生育損傷が緩和された55).これらのことよりHOG経路の活性化は,複合スフィンゴ脂質の生合成の破綻下において出芽酵母の生存率を上昇させる救済経路であることが示された(図4A).このような救済機構としては,TORC2-Ypk1経路を介して,損傷した複合スフィンゴ脂質生合成経路を修復する機構がよく知られていた56, 57).しかしながらHOG経路は,損傷した複合スフィンゴ脂質生合成経路を修復せず,複合スフィンゴ脂質の破綻で二次的に生じる細胞機能異常を補填することがわかった49).Hog1の活性化経路には,形質膜の異常感知センサーであるSln1およびSho1を起点とする経路が知られているが54),複合スフィンゴ脂質の異常に対する救済にはSln1を介した経路が主に機能することがわかった.またHOG経路は,グリセロリン脂質の代謝異常による生育損傷の救済は行わないが,エルゴステロール生合成の異常による生育損傷を緩和することが確認された.このことからHOG経路は脂質マイクロドメインの破綻に対する救済機構であることが想定される49).
高浸透圧ストレス下でHOG経路は,グリセロール-3-リン酸デヒドロゲナーゼの発現上昇を誘導して細胞内グリセロール量を急激に上昇させ,ストレス耐性獲得に寄与することが知られている54).しかしながら,複合スフィンゴ脂質の生合成破綻下でのHOG経路の救済には,グリセロールは寄与していなかった.HOG経路による救済の分子メカニズムを明らかにするために,複合スフィンゴ脂質破綻下でHOG経路依存的に転写応答が起こる遺伝子をトランスクリプトーム解析で調べた結果,59個の遺伝子を見いだした.その中から,複合スフィンゴ脂質の生合成の破綻に対する実質的な救済効果を持つ遺伝子を探索した結果,機能未知の推定Ser/Thrキナーゼ遺伝子がHOG経路下流の救済因子として機能することをつきとめている.本遺伝子の詳細な機能は不明だが,現状では複合スフィンゴ脂質の生合成の破綻によって増加したミトコンドリア由来の活性酸素種の抑制をすることで救済効果を発揮していることが示唆されている.
一方で,プロテインキナーゼA(PKA)の活性化を引き起こす変異(IRA2, PDE2欠損)がAUR1発現抑制下での生育損傷を促進し,逆にPKAの活性を抑える変異(RAS2, GPA2欠損)が生育損傷を抑制することを見いだした(図4B)55).この現象は,PKAが複合スフィンゴ脂質生合成の破綻下で生じるHog1のリン酸化の抑制とMsn2/4の活性化抑制に一部起因することがわかった.HOG経路は,過剰に活性化されるとむしろ細胞毒性を示すことが知られている.そのため,PKAによる抑制は,複合スフィンゴ脂質生合成の破綻下で,HOG経路による救済が細胞毒性に傾かないように適切に調節するために必要であることが考えられる55).
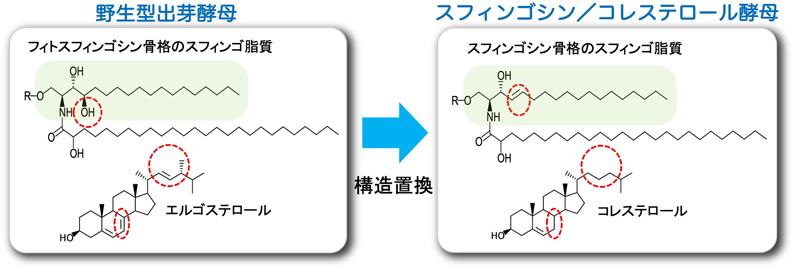
9. 出芽酵母のセラミド構造の哺乳動物型への完全構造置換
異種生物間においてスフィンゴ脂質の基本構造が大きく異なる場合がある.たとえば,哺乳動物における主要複合スフィンゴ脂質であるスフィンゴミエリンは植物や真菌には存在せず,その代わりにIPCが存在する.セラミドの基本骨格となるスフィンゴイド塩基の主要構造は,出芽酵母では水酸化修飾を受けたフィトスフィンゴシン(t18:0)(PHS)であるのに対して,哺乳動物ではトランス二重結合を持ったスフィンゴシン(d18:1)(SPH)である(図5).また,スフィンゴ脂質と機能的連関性が高いとされているステロールについても,出芽酵母ではエルゴステロールなのに対して哺乳動物ではコレステロールとなる(図5).このような構造の違いは,進化の過程で生命がさまざまな機能を獲得したり,環境に適応しようとしたりする中で獲得されたものであると想定されるが,詳細はまったく不明である.そこで我々は,出芽酵母のスフィンゴ脂質のスフィンゴイド塩基の構造をPHSからSPHへと完全に入れ替える(構造置換する)ことによって,どのような表現型が出現するのかを最初に調べることにした58).具体的な方法としては,出芽酵母のSPT遺伝子を欠損させ(lcb2∆),細胞外から任意のスフィンゴイド塩基を供給することでセラミド,複合スフィンゴ脂質の生合成を補完する系を構築した.SPHを供給すると,SPH 1-リン酸が細胞内に蓄積することで毒性が生じるため,LCBキナーゼ遺伝子(LCB4)も欠損させた(lcb2∆ lcb4∆).この変異株は,スフィンゴイド塩基を供給しないと生育できないが,PHSもしくはSPHを供給することで生育が確認された.LC-MS/MSによる解析の結果,SPHを供給した変異株では,SPH骨格を持ったセラミドのみが確認され,さらに複合スフィンゴ脂質の生合成も確認された58, 59).このことは,スフィンゴ脂質のスフィンゴイド塩基構造が哺乳動物型になっても出芽酵母は生育が可能であることを示している.SPHを供給した出芽酵母(SPH酵母)は,PHSを供給した出芽酵母(PHS酵母)と比較して,多面的な環境ストレスに対する耐性能が大きく低下しており,形質膜の透過性亢進,流動性の変化も観察された.またSPH酵母は,細胞壁に損傷をもたらすカルコフロールホワイトに対して高感受性を示し,形質膜と細胞壁の両方に異常が生じていることが示唆された59).
SPH酵母での表現型は,複合スフィンゴ脂質と協調的にマイクロドメイン形成に寄与するステロールが出芽酵母型のエルゴステロールになっていることで引き起こされる可能性が考えられた.そこで,スフィンゴイド塩基と同時にステロール構造も哺乳動物型(コレステロール)に構造置換することを試みた59).ステロール構造置換は,内在性のエルゴステロール生合成を,スクアレンエポキシダーゼをコードするERG1を欠損させることで遮断し,さらに細胞外からのステロール取り込みを行わせるためにステロール生合成遺伝子の発現を制御するUPC2遺伝子の変異体(upc2-1)を発現させることで行った(upc2-1 erg1∆)60).スフィンゴイド塩基とステロールの両方を構造置換できる変異株を用いて,SPH型のスフィンゴ脂質とコレステロールを持った出芽酵母株(SPH/Chol株)を樹立した.SPH/Chol株の複合スフィンゴ脂質量,ステロール量は,野生型と同程度であることが確認された.SPH酵母で観察された多面的ストレス高感受性や形質膜,細胞壁インテグリティの低下は,SPH/Chol株でも同様に観察された.つまり,SPH酵母でみられた表現型がステロールとスフィンゴ脂質の構造的相性に起因しないことが考えられた.また,SPH/Chol株では,出芽酵母の典型的なマイクロドメインの一つであるMCC(membrane compartment of Can1)/エイソソーム61)の数が増大することがわかった.エイソソームは,形質膜ストレスによってその分布パターンを変化させることが知られており62),SPH/Chol株におけるエイソソームの異常は形質膜の性質が変化していることを反映している可能性がある.以上の結果より,出芽酵母のスフィンゴイド塩基,ステロールの構造を哺乳動物型にすると形質膜,細胞壁を含む細胞表面の環境に異常が生じることで,ストレスに対する耐性能が低下してしまうことが示された59).
一般的に生体膜における複合スフィンゴ脂質の存在量は,全体の膜脂質の1–2割程度とされている.出芽酵母における複合スフィンゴ脂質の構造多様性の崩壊もしくは異種生物型への構造置換が,形質膜全体の物性にどのようにして影響を与えるかについては,今後の重要な課題である.本稿で紹介した研究結果より,出芽酵母において複合スフィンゴ脂質は環境ストレスに対する適応に非常に重要であることが示唆されている.一方で,複合スフィンゴ脂質の量的,構造的破綻に対して,典型的なストレス応答経路であるHOG経路,CWI経路,Msn2/4を介した転写応答が救済機構として働くことも見いだしている.これらのことは,出芽酵母における複合スフィンゴ脂質と環境ストレス応答との密接な連関性を示しており,出芽酵母が複合スフィンゴ脂質とストレス応答系の相互関係を通して,さまざまな環境ストレスに対する防御体制をより盤石なものにしようとしている,という考えに帰結する.また,複合スフィンゴ脂質の組成パターンを人為的に改変することで,特定のストレスに対する耐性能を強化した出芽酵母を創成できれば,発酵業界において活躍する出芽酵母に新たな実用性を付与できる可能性も期待できる.
謝辞Acknowledgments
本稿で紹介した研究内容の一部は,前所属である九州大学大学院理学研究院化学部門生体情報化学研究室の大学院生の地道な実験の積み重ねの成果であり,努力の賜物でもあります.また,同研究室の久下理教授(現名誉教授)には多大なご助言をいただきました.出芽酵母のスフィンゴ脂質の哺乳動物型への構造置換の実験の一部は,北海道大学大学院薬学研究の五十嵐靖之教授(現名誉教授),木原章雄教授のご指導のもと行われました.本研究に関わったすべての方々に,この場を借りて厚く御礼申し上げます.
引用文献References
1) Buede, R., Rinker-Schaffer, C., Pinto, W.J., Lester, R.L., & Dickson, R.C. (1991) Cloning and characterization of LCB1, a Saccharomyces gene required for biosynthesis of the long-chain base component of sphingolipids. J. Bacteriol., 173, 4325–4332.
2) Beeler, T., Bacikova, D., Gable, K., Hopkins, L., Johnson, C., Slife, H., & Dunn, T. (1998) The Saccharomyces cerevisiae TSC10/YBR265w gene encoding 3-ketosphinganine reductase is identified in a screen for temperature-sensitive suppressors of the Ca2+-sensitive csg2Delta mutant. J. Biol. Chem., 273, 30688–30694.
3) Guillas, I., Kirchman, P.A., Chuard, R., Pfefferli, M., Jiang, J.C., Jazwinski, S.M., & Conzelmann, A. (2001) C26-CoA-dependent ceramide synthesis of Saccharomyces cerevisiae is operated by Lag1p and Lac1p. EMBO J., 20, 2655–2665.
4) Breslow, D.K., Collins, S.R., Bodenmiller, B., Aebersold, R., Simons, K., Shevchenko, A., Ejsing, C.S., & Weissman, J.S. (2010) Orm family proteins mediate sphingolipid homeostasis. Nature, 463, 1048–1053.
5) Saba, J.D., Nara, F., Bielawska, A., Garrett, S., & Hannun, Y.A. (1997) The BST1 gene of Saccharomyces cerevisiae is the sphingosine-1-phosphate lyase. J. Biol. Chem., 272, 26087–26090.
6) Nagiec, M.M., Skrzypek, M., Nagiec, E.E., Lester, R.L., & Dickson, R.C. (1998) The LCB4 (YOR171c) and LCB5 (YLR260w) genes of Saccharomyces encode sphingoid long chain base kinases. J. Biol. Chem., 273, 19437–19442.
7) Mao, C., Xu, R., Bielawska, A., & Obeid, L.M. (2000) Cloning of an alkaline ceramidase from Saccharomyces cerevisiae. An enzyme with reverse (CoA-independent) ceramide synthase activity. J. Biol. Chem., 275, 6876–6884.
8) Nakahara, K., Ohkuni, A., Kitamura, T., Abe, K., Naganuma, T., Ohno, Y., Zoeller, R.A., & Kihara, A. (2012) The Sjögren-Larsson syndrome gene encodes a hexadecenal dehydrogenase of the sphingosine 1-phosphate degradation pathway. Mol. Cell, 46, 461–471.
9) Ishibashi, Y., Ikeda, K., Sakaguchi, K., Okino, N., Taguchi, R., & Ito, M. (2012) Quality control of fungus-specific glucosylceramide in Cryptococcus neoformans by endoglycoceramidase-related protein 1 (EGCrP1). J. Biol. Chem., 287, 368–381.
10) Ternes, P., Wobbe, T., Schwarz, M., Albrecht, S., Feussner, K., Riezman, I., Cregg, J.M., Heinz, E., Riezman, H., Feussner, I., et al. (2011) Two pathways of sphingolipid biosynthesis are separated in the yeast Pichia pastoris. J. Biol. Chem., 286, 11401–11414.
11) Oura, T. & Kajiwara, S. (2010) Candida albicans sphingolipid C9-methyltransferase is involved in hyphal elongation. Microbiology (Reading), 156, 1234–1243.
12) Takahashi, K., Izumi, K., Nakahata, E., Hirata, M., Sawada, K., Tsuge, K., Nagao, K., & Kitagaki, H. (2014) Quantitation and structural determination of glucosylceramides contained in sake lees. J. Oleo Sci., 63, 15–23.
13) Leipelt, M., Warnecke, D., Zahringer, U., Ott, C., Muller, F., Hube, B., & Heinz, E. (2001) Glucosylceramide synthases, a gene family responsible for the biosynthesis of glucosphingolipids in animals, plants, and fungi. J. Biol. Chem., 276, 33621–33629.
14) Sawada, K., Sato, T., Hamajima, H., Jayakody, L.N., Hirata, M., Yamashiro, M., Tajima, M., Mitsutake, S., Nagao, K., Tsuge, K., et al. (2015) Glucosylceramide contained in Koji mold-cultured cereal confers membrane and flavor modification and stress tolerance to Saccharomyces cerevisiae during coculture fermentation. Appl. Environ. Microbiol., 81, 3688–3698.
15) Saito, K., Takakuwa, N., Ohnishi, M., & Oda, Y. (2006) Presence of glucosylceramide in yeast and its relation to alkali tolerance of yeast. Appl. Microbiol. Biotechnol., 71, 515–521.
16) Rittershaus, P.C., Kechichian, T.B., Allegood, J.C., Merrill, A.H. Jr., Hennig, M., Luberto, C., & Del Poeta, M. (2006) Glucosylceramide synthase is an essential regulator of pathogenicity of Cryptococcus neoformans. J. Clin. Invest., 116, 1651–1659.
17) Nakase, M., Tani, M., Morita, T., Kitamoto, H.K., Kashiwazaki, J., Nakamura, T., Hosomi, A., Tanaka, N., & Takegawa, K. (2010) Mannosylinositol phosphorylceramide is a major sphingolipid component and is required for proper localization of plasma-membrane proteins in Schizosaccharomyces pombe. J. Cell Sci., 123, 1578–1587.
18) Nakase, M., Tani, M., & Takegawa, K. (2012) Expression of budding yeast IPT1 produces mannosyldiinositol phosphorylceramide in fission yeast and inhibits cell growth. Microbiology (Reading), 158, 1219–1228.
19) Tani, M. (2016) Structure–function relationship of complex Sphingolipids in yeast. Trends Glycosci. Glycotechnol., 28, E109–E116.
20) Beeler, T.J., Fu, D., Rivera, J., Monaghan, E., Gable, K., & Dunn, T.M. (1997) SUR1 (CSG1/BCL21), a gene necessary for growth of Saccharomyces cerevisiae in the presence of high Ca2+ concentrations at 37 degrees C, is required for mannosylation of inositolphosphorylceramide. Mol. Gen. Genet., 255, 570–579.
21) Funato, K. & Riezman, H. (2001) Vesicular and nonvesicular transport of ceramide from ER to the Golgi apparatus in yeast. J. Cell Biol., 155, 949–959.
22) Liu, L.K., Choudhary, V., Toulmay, A., & Prinz, W.A. (2017) An inducible ER-Golgi tether facilitates ceramide transport to alleviate lipotoxicity. J. Cell Biol., 216, 131–147.
23) Ikeda, A., Schlarmann, P., Kurokawa, K., Nakano, A., Riezman, H., & Funato, K. (2020) Tricalbins are required for non-vesicular ceramide transport at ER-Golgi contacts and modulate lipid droplet Biogenesis. iScience, 23, 101603.
24) Limar, S., Korner, C., Martinez-Montanes, F., Stancheva, V.G., Wolf, V.N., Walter, S., Miller, E.A., Ejsing, C.S., Galassi, V.V., & Frohlich, F. (2023) Yeast Svf1 binds ceramides and contributes to sphingolipid metabolism at the ER cis-Golgi interface. J. Cell Biol., 222, e202109162.
25) Nagiec, M.M., Nagiec, E.E., Baltisberger, J.A., Wells, G.B., Lester, R.L., & Dickson, R.C. (1997) Sphingolipid synthesis as a target for antifungal drugs. Complementation of the inositol phosphorylceramide synthase defect in a mutant strain of Saccharomyces cerevisiae by the AUR1 gene. J. Biol. Chem., 272, 9809–9817.
26) Uemura, S., Kihara, A., Inokuchi, J., & Igarashi, Y. (2003) Csg1p and newly identified Csh1p function in mannosylinositol phosphorylceramide synthesis by interacting with Csg2p. J. Biol. Chem., 278, 45049–45055.
27) Dickson, R.C., Nagiec, E.E., Wells, G.B., Nagiec, M.M., & Lester, R.L. (1997) Synthesis of mannose-(inositol-P)2-ceramide, the major sphingolipid in Saccharomyces cerevisiae, requires the IPT1 (YDR072c) gene. J. Biol. Chem., 272, 29620–29625.
28) Sato, K., Noda, Y., & Yoda, K. (2009) Kei1: A novel subunit of inositolphosphorylceramide synthase, essential for its enzyme activity and Golgi localization. Mol. Biol. Cell, 20, 4444–4457.
29) Tani, M. & Kuge, O. (2010) Requirement of a specific group of sphingolipid-metabolizing enzyme for growth of yeast Saccharomyces cerevisiae under impaired metabolism of glycerophospholipids. Mol. Microbiol., 78, 395–413.
30) Rivas, M.P., Kearns, B.G., Xie, Z., Guo, S., Sekar, M.C., Hosaka, K., Kagiwada, S., York, J.D., & Bankaitis, V.A. (1999) Pleiotropic alterations in lipid metabolism in yeast sac1 mutants: Relationship to “bypass Sec14p” and inositol auxotrophy. Mol. Biol. Cell, 10, 2235–2250.
31) Tani, M. & Kuge, O. (2014) Involvement of Sac1 phosphoinositide phosphatase in the metabolism of phosphatidylserine in the yeast Saccharomyces cerevisiae. Yeast, 31, 145–158.
32) Moser von Filseck, J., Copic, A., Delfosse, V., Vanni, S., Jackson, C.L., Bourguet, W., & Drin, G. (2015) INTRACELLULAR TRANSPORT. Phosphatidylserine transport by ORP/Osh proteins is driven by phosphatidylinositol 4-phosphate. Science, 349, 432–436.
33) Tani, M. & Kuge, O. (2012) Involvement of complex sphingolipids and phosphatidylserine in endosomal trafficking in yeast Saccharomyces cerevisiae. Mol. Microbiol., 86, 1262–1280.
34) Beeler, T., Gable, K., Zhao, C., & Dunn, T. (1994) A novel protein, CSG2p, is required for Ca2+ regulation in Saccharomyces cerevisiae. J. Biol. Chem., 269, 7279–7284.
35) Zhao, C., Beeler, T., & Dunn, T. (1994) Suppressors of the Ca(2+)-sensitive yeast mutant (csg2) identify genes involved in sphingolipid biosynthesis. Cloning and characterization of SCS1, a gene required for serine palmitoyltransferase activity. J. Biol. Chem., 269, 21480–21488.
36) Yamagata, M., Obara, K., & Kihara, A. (2013) Unperverted synthesis of complex sphingolipids is essential for cell survival under nitrogen starvation. Genes Cells, 18, 650–659.
37) Knupp, J., Martinez-Montanes, F., Van Den Bergh, F., Cottier, S., Schneiter, R., Beard, D., & Chang, A. (2017) Sphingolipid accumulation causes mitochondrial dysregulation and cell death. Cell Death Differ., 24, 2044–2053.
38) Tong, A.H. & Boone, C. (2006) Synthetic genetic array analysis in Saccharomyces cerevisiae. Methods Mol. Biol., 313, 171–192.
39) Morimoto, Y. & Tani, M. (2015) Synthesis of mannosylinositol phosphorylceramides is involved in maintenance of cell integrity of yeast Saccharomyces cerevisiae. Mol. Microbiol., 95, 706–722.
40) Tanaka, S. & Tani, M. (2018) Mannosylinositol phosphorylceramides and ergosterol coodinately maintain cell wall integrity in the yeast Saccharomyces cerevisiae. FEBS J., 285, 2405–2427.
41) Willet, A.H., Wos, M., Igarashi, M.G., Ren, L., Turner, L.A., & Gould, K.L. (2023) Elevated levels of sphingolipid MIPC in the plasma membrane disrupt the coordination of cell growth with cell wall formation in fission yeast. PLoS Genet., 19, e1010987.
42) Koga, A., Takayama, C., Ishibashi, Y., Kono, Y., Matsuzaki, M., & Tani, M. (2022) Loss of tolerance to multiple environmental stresses due to limitation of structural diversity of complex sphingolipids. Mol. Biol. Cell, 33, ar105.
43) Otsu, M., Toume, M., Yamaguchi, Y., & Tani, M. (2020) Proper regulation of inositolphosphorylceramide levels is required for acquirement of low pH resistance in budding yeast. Sci. Rep., 10, 10792.
44) Gatta, A.T., Wong, L.H., Sere, Y.Y., Calderon-Norena, D.M., Cockcroft, S., Menon, A.K., & Levine, T.P. (2015) A new family of StART domain proteins at membrane contact sites has a role in ER-PM sterol transport. eLife, 4, e07253.
45) Li, S.C. & Kane, P.M. (2009) The yeast lysosome-like vacuole: Endpoint and crossroads. Biochim. Biophys. Acta Mol. Cell Res., 1793, 650–663.
46) Tani, M. & Toume, M. (2015) Alteration of complex sphingolipid composition and its physiological significance in yeast Saccharomyces cerevisiae lacking vacuolar ATPase. Microbiology (Reading), 161, 2369–2383.
47) Balguerie, A., Bagnat, M., Bonneu, M., Aigle, M., & Breton, A.M. (2002) Rvs161p and sphingolipids are required for actin repolarization following salt stress. Eukaryot. Cell, 1, 1021–1031.
48) Toume, M. & Tani, M. (2016) Yeast lacking the amphiphysin family protein Rvs167 is sensitive to disruptions in sphingolipid levels. FEBS J., 283, 2911–2928.
49) Yamaguchi, Y., Katsuki, Y., Tanaka, S., Kawaguchi, R., Denda, H., Ikeda, T., Funato, K., & Tani, M. (2018) Protective role of the HOG pathway against the growth defect caused by impaired biosynthesis of complex sphingolipids in yeast Saccharomyces cerevisiae. Mol. Microbiol., 107, 363–386.
50) Chi, Y., Huddleston, M.J., Zhang, X., Young, R.A., Annan, R.S., Carr, S.A., & Deshaies, R.J. (2001) Negative regulation of Gcn4 and Msn2 transcription factors by Srb10 cyclin-dependent kinase. Genes Dev., 15, 1078–1092.
51) Schmitz, H.P., Jendretzki, A., Wittland, J., Wiechert, J., & Heinisch, J.J. (2015) Identification of Dck1 and Lmo1 as upstream regulators of the small GTPase Rho5 in Saccharomyces cerevisiae. Mol. Microbiol., 96, 306–324.
52) Annan, R.B., Wu, C., Waller, D.D., Whiteway, M., & Thomas, D.Y. (2008) Rho5p is involved in mediating the osmotic stress response in Saccharomyces cerevisiae, and its activity is regulated via Msi1p and Npr1p by phosphorylation and ubiquitination. Eukaryot. Cell, 7, 1441–1449.
53) Tanigawa, M., Kihara, A., Terashima, M., Takahara, T., & Maeda, T. (2012) Sphingolipids regulate the yeast high-osmolarity glycerol response pathway. Mol. Cell. Biol., 32, 2861–2870.
54) Brewster, J.L. & Gustin, M.C. (2014) Hog1: 20 years of discovery and impact. Sci. Signal., 7, re7.
55) Urita, A., Ishibashi, Y., Kawaguchi, R., Yanase, Y., & Tani, M. (2022) Crosstalk between protein kinase A and the HOG pathway under impaired biosynthesis of complex sphingolipids in budding yeast. FEBS J., 289, 766–786.
56) Berchtold, D., Piccolis, M., Chiaruttini, N., Riezman, I., Riezman, H., Roux, A., Walther, T.C., & Loewith, R. (2012) Plasma membrane stress induces relocalization of Slm proteins and activation of TORC2 to promote sphingolipid synthesis. Nat. Cell Biol., 14, 542–547.
57) Roelants, F.M., Leskoske, K.L., Martinez Marshall, M.N., Locke, M.N., & Thorner, J. (2017) The TORC2-Dependent Signaling Network in the yeast Saccharomyces cerevisiae. Biomolecules, 7, 66.
58) Tani, M., Kihara, A., & Igarashi, Y. (2006) Rescue of cell growth by sphingosine with disruption of lipid microdomain formation in Saccharomyces cerevisiae deficient in sphingolipid biosynthesis. Biochem. J., 394, 237–242.
59) Kono, Y., Ishibashi, Y., Fukuda, S., Higuchi, T., & Tani, M. (2023) Simultaneous structural replacement of the sphingoid long-chain base and sterol in budding yeast. FEBS J., 290, 5605–5627.
60) Li, Y. & Prinz, W.A. (2004) ATP-binding cassette (ABC) transporters mediate nonvesicular, raft-modulated sterol movement from the plasma membrane to the endoplasmic reticulum. J. Biol. Chem., 279, 45226–45234.
61) Sakata, K.T., Hashii, K., Yoshizawa, K., Tahara, Y.O., Yae, K., Tsuda, R., Tanaka, N., Maeda, T., Miyata, M., & Tabuchi, M. (2022) Coordinated regulation of TORC2 signaling by MCC/eisosome-associated proteins, Pil1 and tetraspan membrane proteins during the stress response. Mol. Microbiol., 117, 1227–1244.
62) Dupont, S., Beney, L., Ritt, J.F., Lherminier, J., & Gervais, P. (2010) Lateral reorganization of plasma membrane is involved in the yeast resistance to severe dehydration. Biochim. Biophys. Acta Biomembr., 1798, 975–985.
著者紹介Author Profile
 谷 元洋(たに もとひろ)
谷 元洋(たに もとひろ)岐阜大学応用生物科学部 教授.博士(農学).
略歴1973年京都府に生る.2003年九州大学大学院生物資源環境科学府博士課程修了.01年日本学術振興会特別研究員(DC2).03年日本学術振興会特別研究員(PD).06年日本学術振興会海外特別研究員.07年九州大学SSP学術研究員.11年九州大学大学院理学研究院化学部門准教授,23年10月より現職.
研究テーマと抱負「スフィンクスの謎」とも例えられている生体膜スフィンゴ脂質の生理機能の解明.出芽酵母の分子遺伝学を駆使することで独自の方向性からスフィンゴ脂質の構造多様性の生物学的意義の解明を目指したい.
ウェブサイトhttps://www1.gifu-u.ac.jp/~sphingo-tani/
趣味楽器演奏,史跡探訪.