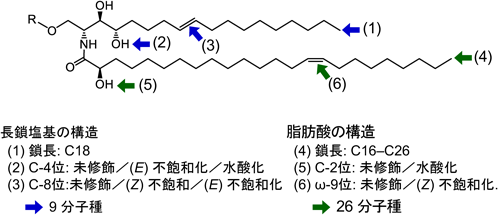
植物のセラミドは,長鎖塩基と脂肪酸の炭化水素鎖中にヒドロキシ化や不飽和化といった複数の化学修飾を受け,位置異性体や幾何異性体を含む複雑な構造多様性が生じている.図1に,植物に普遍的にみられるセラミド構造のバリエーションを示す.長鎖塩基の鎖長はC18のみだが,C-4位とC-8位に3通りずつの修飾パターンがあり,その全組合わせに相当する9種類の分子種が植物体内に広く見いだされる.長鎖塩基C-4位は,真核生物に一般的な不飽和化,もしくはヒドロキシ化を受ける.このうち,植物ではC-4ヒドロキシ型の長鎖塩基を主成分とすることが多く,動物のスフィンゴシン(d18:14E)に対して,フィトスフィンゴシン(4-ヒドロキシスフィンガニン,t18:0)と呼ばれるゆえんである1).植物におけるΔ4不飽和化は,動物のDES1およびDES2と相同な酵素によるが,動物ではDES2がC-4ヒドロキシ化も触媒するのに対し,植物ではヒドロキシ化を担うSBH/DSHファミリーが別に存在する2–4).このファミリーは酵母Sur2のオルソログと考えられるが,酵母では生存に必須ではないのに対し,植物のSBH/DSHの欠損は致死的である4).一方,長鎖塩基Δ4不飽和化の植物における機能はいまだによくわかっていない.菌類では,Δ4不飽和化はΔ8不飽和化やC-9メチル化,およびそれらに続くグルコシルセラミド(glucosylceramide:GlcCer)の生合成に必須であるが5),植物ではΔ4不飽和結合はΔ8不飽和化やGlcCer合成には必須でなく,DESを欠損した植物は大きな異常を示さない6, 7).さらに,アブラナ科など,遺伝子レベルではDESを持つものの,もはやほとんどの組織で発現していない植物もみられる.長鎖塩基Δ4不飽和結合は真核生物に最も普遍的なセラミド構造であるが,植物に限っては進化的に失われていく過程にあるのかもしれない.
長鎖塩基Δ8位の不飽和結合には,Z体とE体の立体異性が存在する.この不飽和化はスフィンゴイド長鎖塩基不飽和化酵素(sphingoid long-chain base desaturase:SLD)が担っており,植物では単独の酵素が二つの立体異性体の両方を生成するというユニークな立体特性を持つ.SLDは菌類や藻類などにも保存されているが,それらはすべてE体のみを生成することから,Z体が植物固有の構造である.被子植物のSLDは,分子系統樹上で大きく二つのグループに分類される8, 9).一方は主にZ体を多く生成する分子種であり,単子葉植物および一部の被子植物(マメ科やナス科など)にみられる.他方はE体を優先的に生成するSLDのグループで,被子植物全般に見いだされる.しかしながら,それぞれのグループ内においても,異性体の生成比はSLDの分子種によって幅があり,二つの異性体がどのような機構で生成されるのかは未解明である.少なくとも,それぞれの植物種においてSLDがさまざまな比率で不飽和異性体を生成することによって,植物種間のセラミドの質的多様性が増大する一因となっていると考えられる.脂肪酸の不飽和結合では,E体よりもZ体の方がより効率的に脂質膜の流動性を上昇させることが知られているが,長鎖塩基Δ8位の不飽和結合についても,飽和,E体,Z体の順に流動性が高くなることが,DESやSLDを遺伝的に改変したイネの形質膜の解析から示されている7).このような膜物性に及ぼす影響は生理的にも重要であると考えられ,低温環境や土壌のアルミニウムイオンといった,生体膜の流動性低下を毒性機序とする環境ストレスへの耐性に寄与していることが明らかになっている7–11).
以上の長鎖塩基の基本構造以外では,第三の不飽和結合を含むスフィンガトリエニン(d18:3)が,トウモロコシやコメ由来グルコシルセラミドのマイナー成分として報告されている12).筆者らも,単子葉類イネ科植物に加え,基部被子植物や裸子植物などで広くスフィンガトリエニンが含まれることを確認しており,中には長鎖塩基総量の1割程度に達するケースも見いだしている.Δ4E,8E,10E-スフィンガトリエニンを有する珪藻やフザリウム菌では,C-10位を不飽和化する酵素遺伝子が同定されている13, 14).しかしながら,植物のゲノムにはこれらに相同性な遺伝子は見いだされないため,未同定の酵素が存在するものと考えられる.また,上記の植物内在性スフィンガトリエニンの検出例はいずれも質量分析によるものであり,不飽和結合の位置および立体異性も未確定である.しかし,SLDを過剰発現させた植物でスフィンガトリエニン含量が増加することがあり,これはZ体を優先的に生成するSLD分子種を発現させた場合に限定されること,また他の長鎖塩基成分と異なり,立体異性体に起因するピーク分離がクロマトグラム上でみられないことなどから,三つの不飽和結合のうち二つはΔ4EとΔ8Zであると推定している.
植物セラミドに含まれる脂肪酸は,鎖長がC16からC26の範囲で,C23やC25などの奇数鎖も比較的多く含まれる.α-ヒドロキシ脂肪酸が主成分で,未修飾のノンヒドロキシ脂肪酸は培養細胞などを除き微量にしか含まれないことが多い.このα-ヒドロキシ基は動物や酵母と相同な酵素であるFAH(fatty acid 2-hydroxylase)が導入しており,水素結合の形成により脂質ラフト様の微小膜脂質ドメインの安定化に寄与していることが,植物細胞のライブイメージングや分子動力学によるシミュレーションから支持されている15, 16).このようなスフィンゴ脂質に依存的な形質膜ドメイン領域には,病原体に対する免疫反応や外因性ストレス依存的なプログラム細胞死の誘導を制御するタンパク質が集積しており,形質膜を介した環境情報伝達に寄与している15–17).また,植物セラミド中の脂肪酸はω9位に不飽和結合を一つだけ含むことがあり,C24以上の極長鎖選択的なアシルCoA不飽和化酵素が同定されている18).このアシル不飽和結合は低温環境での生育に適した植物種で見いだされることが多く,長鎖塩基Δ8不飽和結合と同様に,植物の低温馴化能に寄与していることが遺伝学的に示されている19).
植物の複合スフィンゴ脂質には,モノグリコシル型のGlcCerと,イノシトールリン酸およびオリゴ糖鎖を持つグリコシルイノシトールホスホセラミド(glycosylinositolphosphoceramide:GIPC)の二つのクラスが存在する.これらの量比は植物種や組織によって異なるが,おおよそ2 : 1–1 : 5程度であり,両者を合わせると全スフィンゴ脂質の90%ほどを占める.また,これら2種の複合スフィンゴ脂質はセラミド骨格にも明確に異なる特徴を示す.GlcCerにはΔ8Z不飽和型の長鎖塩基と比較的短い脂肪酸(C16–C20)が含まれるが,GIPCにはC-4ヒドロキシ型かつΔ8飽和またはE不飽和型の長鎖塩基と,C24を中心とする極長鎖脂肪酸の組合わせからなるセラミドが選択的に含まれる.それぞれの親水部転移反応に関わる酵素群はセラミド構造に対する厳密な基質特異性を示さないことから,基質となるセラミドを細胞内で区画化することが,このようなセラミド構造の偏りを生む機構として考えられる.植物では,GlcCer合成酵素はセラミド合成系と同じ小胞体に局在し,GIPC合成酵素群はゴルジ体に局在する.そのため,特定のセラミド分子を構造依存的に選択して小胞体からゴルジ体へと輸送する因子が存在すると考えられている.植物にも動物のCERTのようなセラミド輸送体が存在するのか,今後の研究の進展が期待される.
植物の複合スフィンゴ脂質では,長らくGlcCerが研究の中心であった.一方,GIPCについては,1958年にはすでにその存在が知られていたが20),その後スポットライトが当たるようになったのは最近になってからである.2000年代の後半から2010年代にかけて,GIPCを分析するリピドミクス技法が発展したことによって21),構造の多様性や生合成機構など,分子レベルでの理解が急速に進んだ.
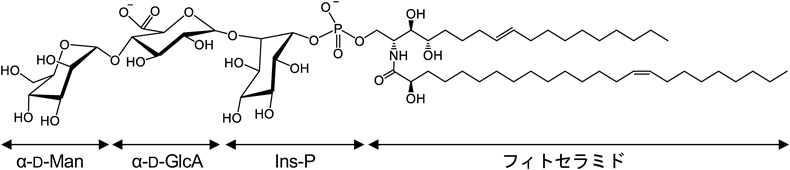
植物GIPCの基本構造を図2に示す.GIPCの親水部新規合成の初発反応は,ホスファチジルイノシトールからセラミドへのイノシトールリン酸の転移反応である22).これは酵母など菌類のイノシトールホスホセラミド(inositolphosphoceramide:IPC)の合成系と同一の反応で,これを触媒する植物のIPCS(inositolphosphoceramide synthase)および酵母のIPCSであるAur1は,互いに,また動物のスフィンゴミエリン合成酵素ともそれぞれわずかに相同性を示す.ちなみに,抗真菌性の抗生物質であるオーレオバシジンAはAur1の阻害剤であり,本薬剤耐性に関わる真菌遺伝子としてAur1は見いだされたのであるが,植物IPCSにはまったく阻害作用を示さない.一方,植物のIPCSを選択的に阻害する化合物もケミカルスクリーニングにより見いだされており,こちらは耐性の生じにくい新規除草剤として期待されている23).
初発反応により生じたイノシトールホスホセラミドを中間体として,さらに糖転移反応が連続的に続く.第一糖にはグルクロン酸が付加される.この反応を触媒する酵素IPUT1(inositolphosphoceramide glucuronosyltransferase 1)は,細胞壁多糖の生合成に関与するグルクロン酸転移酵素ファミリーから遺伝子重複によって分岐派生したと推定される24).細胞外に露出したGIPC糖鎖は,細胞壁を構成する多糖分子と物理的に相互作用していると考えられており25),これらが合成酵素の進化的な面でも近い関係にあることは非常に興味深い.
続いて第二糖には,マンノース(Man)もしくはN-アセチルグルコサミン(GlcNAc)のいずれかが転移される.前者はGMT1(GIPC mannosyltransferase 1),後者はGINT1(glucosamine inositolphosphoceramide transferase 1)と呼ばれる糖転移酵素がそれぞれ担っている26, 27).ここまでが陸上植物におけるGIPCの最小構造であり,これより小さな親水部を持つGIPCは天然に見いだされず,また合成系の遺伝子欠損は致死的である.それぞれの中間体に相当する分子は,合成系の変異体を除き植物体内にほとんど検出されないため,小胞体からゴルジ体へ輸送されたセラミドは,一連の転移反応を連続的に受け,速やかに成熟型GIPCまで合成が進むものと考えられる.
さらに,植物によっては第二糖以降も糖鎖伸長が進む.多くの双子葉植物では第二糖までの最小構造を主成分とするが,イネをはじめとする単子葉植物では,第二糖残基のGlcNAcに続いてヘキソース(分子種未同定)が付加された三糖型GIPCを主成分とする種が多い28).第四糖以降にさらにヘキソースやペントースが複数連なったより大きな糖鎖構造も見いだされているが29),天然存在量が少なく,詳細な結合様式や合成系はよくわかっていない.
GIPCの構造実体や合成系が明らかになってきたことで,機能面についても次第に報告例が蓄積してきている.ここではいくつか代表的な研究例を紹介する.GIPCの第一糖であるグルクロン酸は,そのカルボキシ基にNa+が結合すると,形質膜上で隣接するCa2+チャネルを活性化し塩ストレス応答を誘導する,いわば塩害センサーとしての機能を持つことが提唱されている30).またGIPC糖鎖は,NLP(necrosis and ethylene-inducing peptide 1-like)と呼ばれる植物病原菌に広く保存された細胞壊死性タンパク質の分子標的となっている.二糖型GIPCに結合すると,NLPは構造変化を引き起こして形質膜に小孔を形成し破壊するが,三糖型GIPCに結合した場合には,ヘキソース残基一つ分の差により形質膜を損傷できない.このことが,NLPを持つ病原菌と宿主植物の間の選択性を決定する要因となっている31).
植物GIPCは,第二糖残基の分子種によって,Man型またはGlcNAc型の二つのグループに分類される.それぞれ異なる酵素によって作り分けされているが,その生物学的意義はよくわかっていない.たとえば,双子葉植物のシロイヌナズナは栄養組織の全体にMan型糖鎖のみを持ち,単子葉植物のイネは逆にGlcNAc型糖鎖のみを持つ.これはシロイヌナズナ組織ではGMT1のみが,イネ組織ではGINT1のみが発現していることで説明できるが,それぞれの遺伝子を機能破壊すると,共通の前駆体であるグルクロノシルイノシトールホスホセラミドが蓄積し,いずれの植物も実生致死の表現型を示す27).この結果からは,Man型とGlcNAc型は,植物の生存に必須な機能を同じように担っているようにも思われる.しかしながら,Man型を欠損したシロイヌナズナgmt1変異体に,GINT1を強制発現させることでGlcNAc型糖鎖を合成させた異種糖鎖相補系統は,依然として重篤な生育不全を示す27).このことから,二つの糖鎖型が機能レベルで同等でないことは明らかである.植物GIPCの糖鎖構造の違いがどのような機能的差異を有するのか,また植物種によってこれらの使い分けが異なるのはなぜか,今後の研究で明らかにしていきたいと考えている.
上述の植物GIPCの親水部構造は,コケ植物から種子植物のいわゆる陸上植物に加え,より原始的な車軸藻類まで保存されていることから,植物の陸上化以前には基本的な構造がすでに誕生していたものと考えられる.一方,車軸藻類よりもさらに原始的な藻類におけるスフィンゴ脂質の進化の足跡は,いまだはっきりしていない.最近,植物の進化系統で最も原始的な生物種である紅藻の一種Galdieria sulphurariaにおいて,陸上植物と同様の糖鎖構造を持つGIPCが報告された32).しかしながら,進化上,紅藻と車軸藻の中間に位置する緑藻では,イノシトールリン酸型の複合スフィンゴ脂質を含む種は見つかっているものの,それらの糖鎖構造は植物型とは異なっている29).さらに最近,我々は最小の真核藻類(ピコオルガ)として知られる原始的な緑藻種オストレオコッカス(Ostreococcus tauri)において,IPCおよびGlcCer系列のセラミド種は合成酵素遺伝子のレベルで存在しないこと,代わりに複数のウロン酸とヘキソースで構成されるオリゴ糖鎖が直接セラミドに付加された,新奇の酸性糖脂質クラス(たとえば,Hex-Hex-HexA-HexA-ceramide.Hexはヘキソース,HexAはウロン酸を示す)を含有することを見いだしている33).少なくとも,これらの原始的な藻類では,種や系統ごとに複合スフィンゴ脂質の構造体系が大きく異なっているようである.近年,高速シークエンサーの普及に伴い藻類のゲノム情報の整備が進んでいる.合成系の遺伝子情報と実際の脂質組成を照合しながら,より大規模に複合スフィンゴ脂質の体系分類を行う必要があろう.
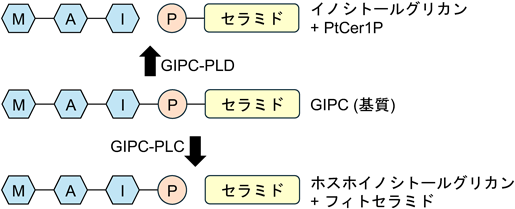
徳島大学の田中らは,キャベツを破砕したホモジネート中に,アルカリ耐性かつリン酸を含有する未知の脂質成分が生じることを見いだし,これがGIPCの分解によって生じたフィトセラミド1-リン酸(phytoceramide 1-phosphate:PtCer1P)であることを突き止めた34).リン脂質のリン酸ジエステル結合を親水部側で切断する,いわゆるホスホリパーゼD(PLD)様の反応であることから,この酵素はGIPC-PLDと命名された.その後,筆者も研究グループに参画し,最近ようやく突き止めることに成功したその酵素実体は,驚くべきことにそれまでグリセロ脂質ホスホリパーゼCとして知られていたNPC(non-specific phospholipase C)であった35).当初,本酵素はダイコン(Raphanus sativus L.)から単離され,他の植物のNPCとの相同性からRsNPC3aと命名された.さらに,シロイヌナズナ(Arabidopsis thaliana L.)の相同因子AtNPC3に加え,NPCファミリーの中で最も近縁なAtNPC4も同様にGIPC-PLD活性を持つことが見いだされた.特にAtNPC4は,先に他のグループによってGIPC分解活性を持つことが報告されていたものの,そこではホスホリパーゼC,すなわち産物としてPtCer1Pではなくフィトセラミドを産生するGIPC-PLCであるとされていた36).しかしながら,我々の実験系においては,先行論文と同様に大腸菌で発現させた3種の組換えタンパク質(RsNPC3a, AtNPC3, AtNPC4)は,いずれもGIPCを基質とした際にPtCer1Pのみを生成し,フィトセラミドはまったく生成しなかった.また,植物組織に一過的もしくは恒常的に過剰発現させた場合にも,同様にPtCer1Pのみを生じたことから,NPCはin vitroとin vivoのいずれにおいてもGIPC-PLDであることが示されている.一方,こうしたGIPC-PLDの研究を進める過程で,植物組織にはGIPCからフィトセラミドを生じるGIPC-PLC活性も共存することがわかってきている37).当初考えられていたNPCは,実際にはGIPC-PLC活性を示さないことから,他に未同定の酵素が存在すると考えられる.
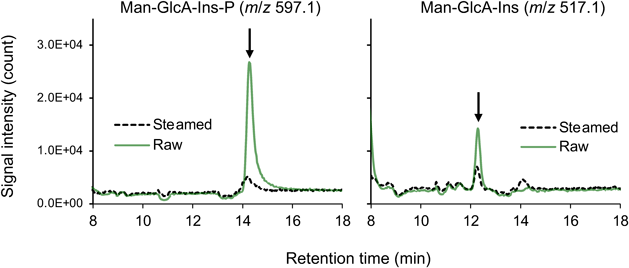
図3に,2種類の酵素によるGIPC分解反応で生じる産物の構造を示す.GIPC分解活性を持つかどうか,またPLDとPLCのいずれの活性を有するかは,植物種によっても異なる.たとえば,アブラナ科に属する植物では両方の酵素活性が検出されることが多いが,キャベツやダイコンではGIPC-PLD活性が強く,シロイヌナズナなどではGIPC-PLC活性が主要である.これらの酵素活性は,通常は疎水部側の産物であるPtCer1PとフィトセラミドをTLCまたはLC-MS/MSで検出することにより評価するが,筆者らは親水部側の反応産物(イノシトールグリカンおよびホスホイノシトールグリカン)をHILIC(hydrophilic interaction liquid chromatography)-MSを用いて検出する系も確立している(図4).疎水部と親水部の両方の分解産物データを合わせて考えると,上述のGIPC分解反応は,末端から糖を削っていくエキソグリコシダーゼ型でなく,リン酸ジエステル結合を切断して親水部と疎水部に切り分けるホスホジエステラーゼ反応によるものであることがわかる.
GIPCの分解反応が見つかったことで,次なる興味はその生物学的な存在意義である.これまでに提唱されている機能として,リン酸欠乏時における貯蔵リンの代謝再分配があげられる.リン酸は代謝におけるエネルギー輸送の担い手であり,植物細胞内のリン酸濃度は光合成炭素同化反応の主要な律速要因の一つである.そのため,リン酸欠乏下におかれた植物では,生体膜を構成していたリン脂質を分解してリン酸を取り出すと同時に,性質の近い糖脂質に置き換える膜脂質転換が速やかに誘導される.シロイヌナズナのAtNPC4は,リン酸欠乏下において高レベルに発現し,GIPCからGlcCerへの膜脂質転換によるリン酸供給に寄与することが報告されている36).一方,AtNPC3のように,リン酸欠乏と遺伝子発現が相関しないものは,他の生理機能へ寄与しているものと推測される.それは今後の研究課題であるが,機能性を考える上で重要なポイントは,GIPCが分解される場所である.通常,GIPCは形質膜の外葉に偏局在している38).動物の形質膜におけるスフィンゴミエリン分解と同様の反応を考えると,生じたフィトセラミドまたはPtCer1Pが形質膜上で脂質ラフトのような膜構造の変化を誘導するか,細胞内に移行して何らかの情報伝達へ寄与することが示唆される.また,スフィンゴミエリンの場合と異なる点として,GIPCからは細胞外マトリクスにイノシトールグリカンが遊離することがあげられる.これは分子量が500を超える特異構造を有する親水性分子であり,細胞間移行性のリガンドとして機能する可能性が考えられる.上述のように,GIPCの親水部には構造多様性が存在するため,その分解によって生じる分子情報にも多様性が生じることになる.現在,筆者らのグループでは,植物におけるGIPC分解系の全容を明らかにすることを目指して,GIPC分解酵素を遺伝的に改変した植物系統や,親水部および疎水部の分解産物を網羅的に分析するメタボロミクス的アプローチを用いた研究を精力的に進めているところである.