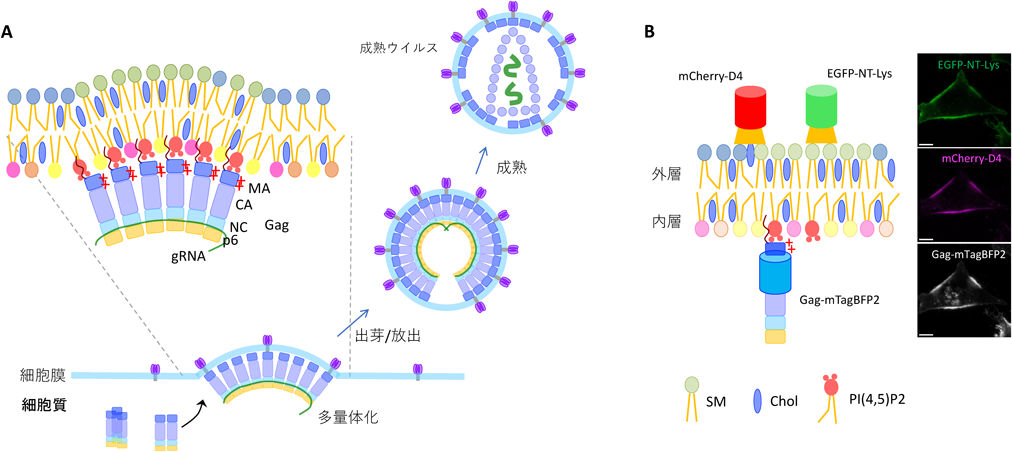
ヒト免疫不全症ウイルス1型(HIV-1)は,エイズの原因ウイルスの一つであり,全世界で3900万人の感染者が報告される(2022年時点)公衆衛生上の脅威である.HIV-1は被膜ウイルスであり,複製サイクル後期に宿主感染細胞の細胞膜から出芽する際に,宿主膜脂質を被膜として獲得する.この一連の過程に中心的な役割を果たすのがウイルス由来のマルチドメインタンパク質,Gag(Group-specific antigen)である.Gagは宿主細胞の細胞質で翻訳されたのち,N末端matrix(MA)ドメインのミリストイル化と塩基性アミノ酸に富んだ配列によって,細胞膜内層の酸性リン脂質PI(4,5)P2にターゲットされる.膜上にリクルートされたGagは,Gag latticeと呼ばれる多量体を形成し,ウイルス粒子形成の足場となる.多量体の形成とともにlattice固有の膜曲率を持った新粒子の出芽が進行し,最終的に宿主タンパク質の働きで細胞外へ放出される1)(図1A).
複数のリピドミクス解析により,ウイルス粒子にはスフィンゴミエリン(SM),コレステロール(Chol)やPI(4,5)P2など,特定の脂質が,宿主細胞の細胞膜に比べて濃縮されていることが報告されている2).
哺乳動物細胞の細胞膜は,構成脂質が内外層で非対称的に分布する特徴的な構造をとる.SMは細胞膜脂質の約10%を占め,その90%が外層に存在する主要な構成脂質である.Cholもまた,細胞膜脂質の約40%を構成する主要な脂質であるが,その層間分布は決定されていない.SMとCholはその高い親和性によって,細胞膜上で集合体(脂質マイクロドメイン)を形成すると考えられている3).ただし,細胞膜における存在比は,CholがSM以外の細胞膜脂質とも相互作用しうることを示唆している.
細胞膜内層で多量体形成するGagが,SMのような外層の脂質を濃縮するメカニズムは不明である.近年,蛍光脂質類似体を用いた解析が行われたが,必ずしも濃縮を説明しない4, 5).そこで筆者らは,内在性の脂質を可視化し,その分布・動態を定量的に評価することでこの問題に取り組んだ6).
2. Gagによる細胞膜脂質ドメイン再編成の可視化
はじめに,細胞膜内層でGagが多量体を形成し,出芽に至る過程における内在性の脂質の分布・動態の可視化を行った.Gagタンパク質の単独発現により,感染・複製能を持たないウイルス様粒子(virus-like particles:VLPs)を産生できることが広く知られており,本研究でもこの実験系を用いた.そしてGagの可視化には,N末端のMAとcapsid(CA)ドメインの間に蛍光タンパク質(FP)を挿入した構築Gag-FPを用いた.また,多量体形成時のGag-FPによる立体障害を避けるために,タグなしGagとGag-FPの3 : 1混合物を発現させた.内在性のSMおよびCholの可視化には,NT-Lys(non-toxic lysenin)およびD4をプローブとして使用した.両者はそれぞれ,ミミズEisenia foetida,細菌Clostridium perfringens由来の孔形成毒素lysenin, perfringolysin Oの毒性のない脂質結合ドメインであり,それぞれ5分子以上のSMの集合体,40%以上のCholを含む膜に特異的に結合する7, 8).そのため本稿では,これらのプローブによって検出された脂質ドメインを,それぞれSMまたはCholに富んだ脂質ドメインと呼ぶ.
実験では,HeLa細胞にGagを一過性に導入し,24時間後に細胞膜外層の脂質を蛍光脂質プローブで標識したサンプルを観察・計測に供した(図1B).
ウイルス粒子は直径100~150 nm,脂質マイクロドメインは直径5~50 nmとされる.このサイズは光学顕微鏡の回折限界以下であるため,観察・計測には超解像顕微鏡や異なるプローブ間の距離や比率に感度の高い,FLIM-FRET(fluorescence lifetime imaging based on Förster resonance energy transfer)を使用した.また,脂質ドメインの動態計測にはFRAP(fluorescence recovery after photobleaching)を使用した.
1)Gagと脂質ドメインの超解像観察
GagがSMとCholをウイルス粒子に濃縮することから,まず細胞膜を挟んだGagと脂質ドメインの共局在を超解像顕微鏡PALM/dSTORM(photoactivated localization microscopy/direct stochastic optical reconstruction microscopy)により観測した.具体的には,Gag/Gag-mEos4bを発現させた細胞の細胞膜外層を,SNAPタグを介してAlexa Fluor 647で標識したAF647-NT-Lys,またはAF647-D4で染色した.mEos4bは,UV域の光照射により蛍光が緑から赤へ変化する(photoconversion),PALMに適したタンパク質である.二量体を形成しうる親mEos2への変異導入により作製されたmEos4bは,ほぼ完全に単量体であり,蛍光特性も改良された.また,Gag多量体化の効果を評価するために,多量体化不全変異体Gag-WM9)を使用した.Mander’s overlap coefficientは,検出された約50%のGagがSMに富んだ,あるいはCholに富んだ脂質ドメインと共局在していることを示した.この共局在は,多量体化によって増加した.
また,超解像顕微鏡によって局在化した脂質プローブのクラスター解析により,ドメインサイズを推定した.SMに富んだ脂質ドメインの直径は,野生型Gagの発現によってのみ有意に増加したが,Cholに富んだ脂質ドメインの直径はGagの発現によって変化しなかった.これらの結果は,Gagは多量体化の際にSMに富んだ脂質ドメインを集め,より大きなドメインを形成することを示唆している.
2)脂質ドメイン動態への影響
さらに,脂質ドメイン動態へのGagの影響を検証するために,EGFP-NT-LysまたはEGFP-D4のFRAPを行った.コントロールではEGFP-NT-Lysの蛍光は85%まで回復した(mobile画分)が,Gag存在下では蛍光回復は64%に抑えられた.すなわち,immobile画分が15%から36%に有意に増加した.この結果は,Gagの発現がSMに富んだ脂質ドメインの拡散を抑えることを示している.一方で,Cholに富んだ脂質ドメインは,Gagの発現にかかわらず常に動きが抑えられていた.
4. 2種類の異なる脂質ドメイン分布へのGagの影響
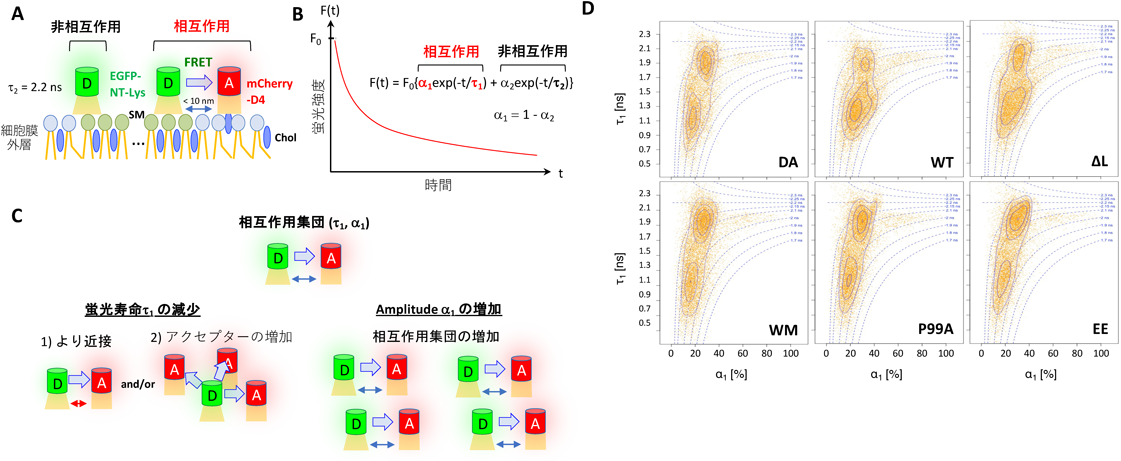
次に,2種類の異なる脂質ドメインの相対的な分布・動態に対するGagの影響を評価するために,アクセプターmCherry-D4存在下,ドナーEGFP-NT-LysのFLIM-FRETを行った.ドナーとアクセプターが存在する条件では,二つのドナー集団が予測される.一方は,アクセプターとFRETを生じる距離にない(非相互作用)集団であり,他方はFRETを生じる距離内に存在する(相互作用)集団である(図2A).各ピクセルにおける蛍光減衰曲線は,これら二つの集団の寄与を含み,二つの指数関数の和でよく近似される.ここでは,τ2をドナーのみで標識したサンプルから得た蛍光寿命に固定して,相互作用集団の蛍光寿命τ1[ns]とそのamplitude α1[%]を得た.また,α1とα2の和は1であり,各ピクセルにおける相互作用集団と非相互作用集団の割合を示す(図2B).本研究では,相互作用集団の分布を2Dグラフで可視化した他,統計検定のために1Dグラフにおいても解析した.本稿では,2Dグラフの結果を紹介する.Gag非存在下(図2D, DA),相互作用集団は二つのピーク[(τ1, α1)=(1.1, 20), (1.9, 30)]を示す集団に分かれた.FLIM-FRETでは,ドナー・アクセプターがより近接する場合や,ドナーと近接するアクセプター数が増える場合に蛍光寿命が減少する(図2C).すなわち,短い蛍光寿命の集団(1.1 ns, 20%)は,長い蛍光寿命の集団(1.9 ns, 30%)に比べ,アクセプターとより近接しているかつ(または)周りのアクセプター数のより多いドナー集団を表している.野生型Gag存在下(図2D, WT),相互作用集団はDA同様に二つのピーク[(τ1, α1)=(1.2, 25), (1.9, 30)]を持つ集団に分かれた.しかし,短い寿命の集団(1.2 ns, 25%)が増加した一方で,長い寿命の集団(1.9 ns, 30%)は減少した.また,短い寿命の集団のamplitudeがDAの20%から25%に増加した.この結果は,Gagが2種類の異なる脂質ドメインを近接・再編するとともに,近接するドメインを増加することを示唆している(図2C).この結果は,AF647-NT-LysとJanelia Fluor 549-D4のdSTORM計測において,両ドメインの共局在がGag発現により増加する結果とも一致した.
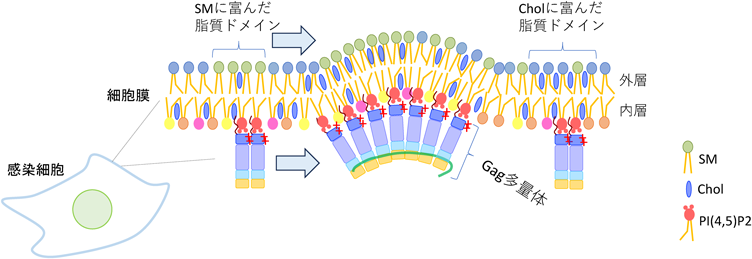
以上より,Gagは細胞膜外層のSMあるいはCholに富んだ脂質ドメインの裏側(内層)に存在するPI(4,5)P2に到達したのち,多量体形成に伴って近傍のSMに富んだ脂質ドメインを捕捉してより大きな脂質ドメインを形成すると推測された(図3).
最後に,多量体化に伴い形成される膜曲率の脂質ドメイン再編への効果を検証した.そのためにGag-∆L, Gag-P99A, Gag-EE変異体を使用してFLIM-FRET実験を行った.P99A, EEは平たい多量体を形成するため小さい膜曲率を示し,∆LはVLPが膜から切り離されないため野生型の膜曲率を持った粒子を膜上に蓄積する4, 10, 11).実験の結果,膜曲率の小さいP99A, EEはGagの発現のないDAや多量体形成不全WMと,膜曲率の大きい∆Lは野生型WTと同様のパターンを示した(図2D).以上の結果は,膜曲率がSMに富んだ脂質ドメインとCholに富んだ脂質ドメインの近接・再編に寄与していることを示している.
なぜウイルスはSMとCholを被膜に濃縮するのだろうか? 生理的温度における水–脂質単成分系では,シス型の不飽和アシル鎖を持つ脂質の二重膜は,アシル鎖の配向が乱れた,流動性の高い,“柔らかい”膜,液体無秩序相(液晶相)[liquid-disordered phase]を示す.一方,SMのようなスフィンゴ脂質など,飽和アシル鎖を持つ脂質の二重膜は,アシル鎖の配向がそろい,非常に緊密に充填された,流動性のないゲル相を示す.ゲル相へのChol添加によりアシル鎖の配向はそろいつつも流動性を保持した,密に充填され,透過性の制限された,“硬い”膜,液体秩序相[liquid-ordered(Lo)phase]が生じる.冒頭でふれたSMとCholからなる脂質マイクロドメインは,Lo phaseの性質を示すと考えられている3).リピドミクスの結果や,環境依存性色素Laurdanを用いた解析12)からウイルス被膜もまたLo膜であると推定されている.CholやSMを除いた宿主細胞膜由来の粒子の感染性が低下するため,脂質マイクロドメインによって付与される被膜の“硬さ”が必要である可能性が指摘されている.
最近,宿主由来の中性スフィンゴミエリナーゼ(nSMase2)が,ウイルス粒子の成熟に必要であることが報告された13, 14).nSMase2は,主に細胞膜内層(細胞質側)に局在して,SMを加水分解してセラミド(Cer)を生じる.Cerは水酸基からなる小さい親水性頭部を持ち,SM同様,飽和アシル鎖を疎水性部分とする,負の膜曲率を持つ脂質である.こうした分子構造に加え,頭部の水酸基を絡めた水素結合ネットワークを構成することにより,Cerはゲル相のドメインを形成する.生体膜においても,SMaseの作用で一過性に生じたドメインが,キナーゼ/ホスファターゼなどをリクルートしてシグナル伝達に機能すると考えられている(Cerドメイン).nSMase2を薬理学的あるいは遺伝学的に阻害した細胞は,形態の異常な,Gagプロセシング不全を示す未成熟で,低感染性の粒子を産生する.現時点では,どのタイミングでnSMase活性が必要なのか,外層のSMがどのように粒子内のnSMaseに提示されるのか等,詳細は不明である.これら形態異常の粒子の電子顕微鏡観察では,Gag latticeのない部分の被膜が外側に突出する.Gag latticeは被膜内層表面を完全には覆わないため,latticeのない部分の被膜の強度と膜曲率を保証するために,“硬い”かつ負の膜曲率を持ちうるCerドメインが必要なのかもしれない.
細胞膜内層と外層の脂質ドメインは膜を挟んで共役していると考えられているが,そのメカニズムは不明である3).人工膜では,Gagは多量体化に伴い蛍光PI(4,5)P2を消光する.すなわち,内層のPI(4,5)P2をクラスター化する結果が得られている15).今回の結果から,Gagがこの表裏の共役を利用していることが推測され,本研究で得られた知見や実験系がその解明に貢献することを期待している.