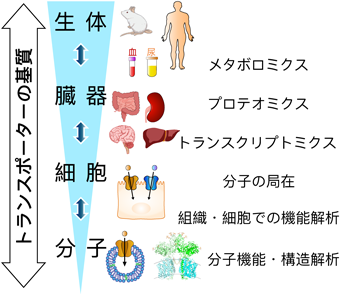
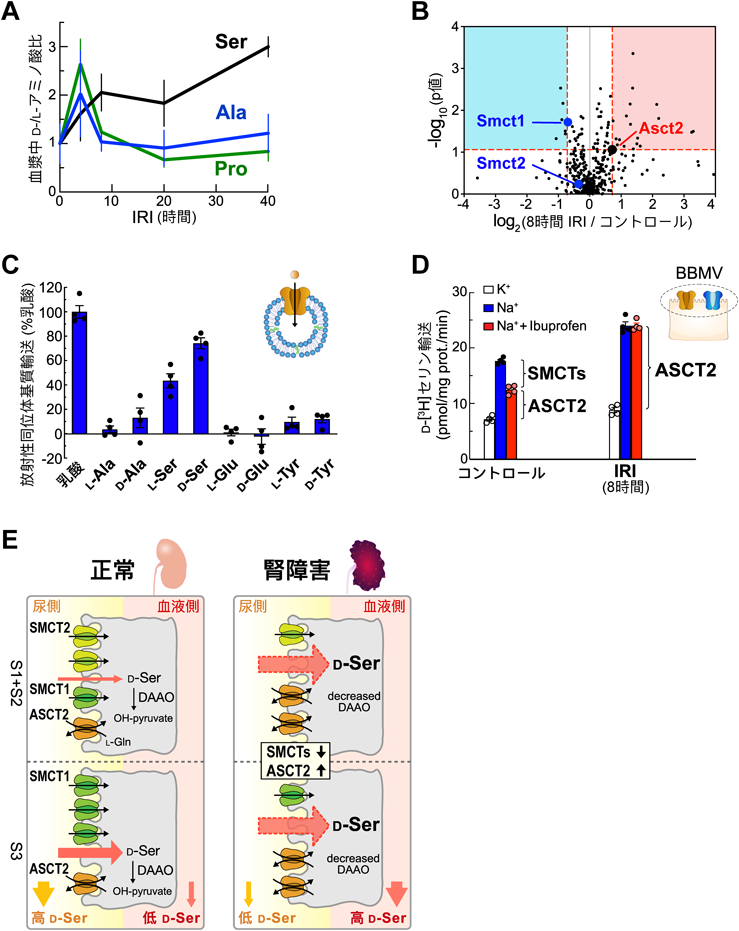
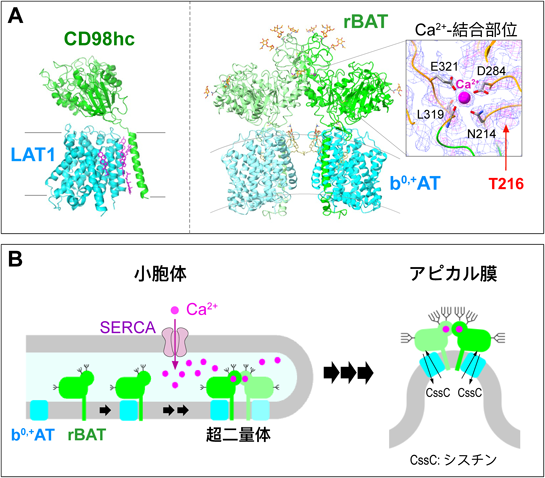
生体における膜輸送体の機能を明らかにする「多階層的アプローチ」A “Multi-hierarchical Approach” to elucidate the functions of membrane transport proteins in living organisms
1 岩手大学農学部応用生物化学科Department of Biological Chemistry and Food Science, Faculty of Agriculture, Iwate University ◇ 〒020–8550 岩手県盛岡市上田3–18–8 ◇ 3–18–8 Ueda, Morioka, Iwate 020–8550, Japan
2 東京慈恵会医科大学SI医学応用研究センターCenter for SI Medical Research, The Jikei University School of Medicine ◇ 〒105–8461 東京都港区西新橋3–25–8 ◇ 3–25–8 Nishi-shimbashi, Minato-ku, Tokyo 105–8461, Japan
3 東京慈恵会医科大学臨床検査医学講座Department of Laboratory Medicine, The Jikei University School of Medicine ◇ 〒105–8461 東京都港区西新橋3–25–8 ◇ 3–25–8 Nishi-shimbashi, Minato-ku, Tokyo 105–8461, Japan
発行日:2024年10月25日Published: October 25, 2024