自然免疫は,病原体に対する初期免疫応答を担う重要なシステムである.ウイルス感染に対して,マクロファージや樹状細胞といった自然免疫細胞は,ウイルス由来の核酸を認識することで,迅速に炎症応答を誘導しさまざまな炎症性メディエーターを産生する.中でも抗ウイルス免疫応答の中心的な役割を果たしているのがI型インターフェロン(IFN)である.I型IFNは,分泌細胞自身や近傍細胞に対しIFN-α/β受容体(IFNAR)を介して細胞内のJAK-STAT経路を活性化し,I型IFN誘導性遺伝子群(IFN-stimulated genes:ISGs)の発現を誘導する.ISGは,ウイルスゲノムの複製を抑制する,あるいは感染細胞に対して細胞死を誘導することで,抗ウイルス作用を発揮する.このような一連の抗ウイルス応答は,Toll様受容体(TLR)やretinoic acid-inducible gene I(RIG-I)様受容体(RLR)に属するRNAセンサー分子や,cyclic GMP-AMP synthase(cGAS)と呼ばれるDNAセンサー分子によって,ウイルス核酸を直接認識することで惹起される1).
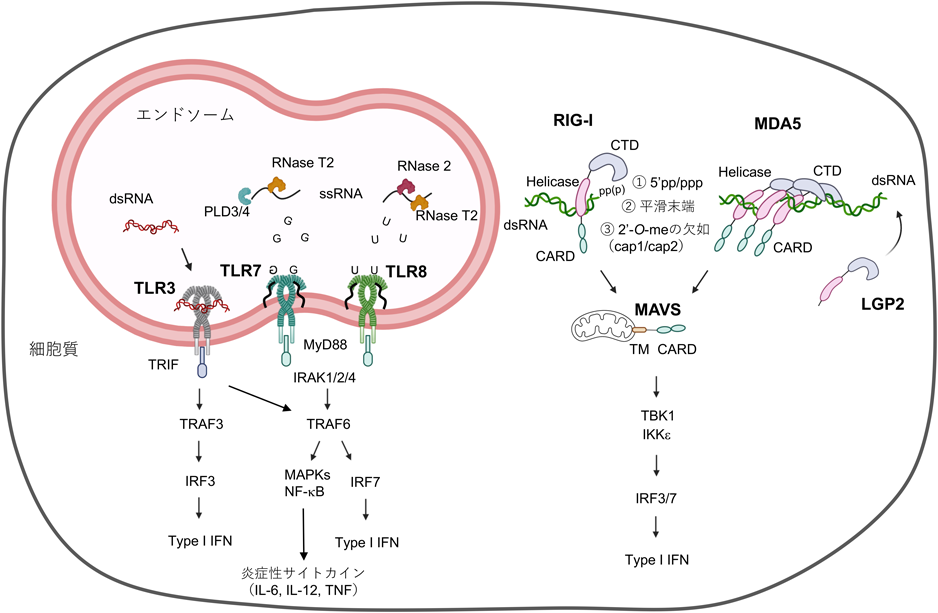
核酸を認識するTLR(TLR3, TLR7, TLR8, TLR9, TLR13)はエンドソームに局在しており,貪食により捕捉された病原体から放出されるリガンドを認識し,病原体排除に重要な役割を持つ2, 3)(図1).TLRシグナルの伝達には,細胞内ドメインに高度に保存されたToll-interleukin(IL)-1 receptor(TIR)ドメインが下流のアダプター分子と相互作用することが必要である.TLR3を除くTLRはすべて,MyD88と呼ばれるアダプタータンパク質がTIRドメインを介してTLRと会合し,下流のシグナルを活性化させる.MyD88はIL-1 receptor–associated kinase(IRAK)-1/2/4を介してTNF receptor-associated factor 6(TRAF6)をリクルートし,K63型ポリユビキチン鎖の生成を介してTGF-β-activated kinase1(TAK1)を活性化する.続いてIκB kinase(IKK)複合体やmitogen-activated protein kinase(MAPK)といったシグナルモジュールを介してNF-κBやAP-1などの転写因子を活性化する.一方,TLR3はTIR domain-containing adaptor inducing IFN-β(TRIF)と呼ばれるアダプタータンパク質がTIRドメインを介して相互作用する.TRAF3やTANK-binding kinase 1(TBK1)をリクルートし,転写因子であるinterferon regulatory factor 3(IRF3)を活性化することでI型IFNを誘導する.また同時にMAPKシグナルの活性化やNF-κBによる炎症性サイトカインの誘導も引き起こす.
RLRファミリーに属するRIG-Iとmelanoma differentiation-associated protein 5(MDA5)は,ほぼすべての有核細胞に発現し,細胞質内においてそれぞれ長さの異なる二本鎖RNAを認識しI型IFN応答を引き起こす4)(図1).RIG-IとMDA5はともに,N末端に二つのcaspase activation and recruitment domain(CARD),DExD/H box型RNAヘリカーゼ,さらにRNA認識に重要なC末端ドメインを持つ.二本鎖RNAを認識するとRIG-IやMDA5は,ユビキチン化やリン酸化など翻訳後修飾を受け,ミトコンドリア上でmitochondrial antiviral-signaling protein(MAVS)とCARDドメインを介した相互作用により重合化が起こる.続いてMAVSはTBK1-IκB kinase-ε(IKKε)を活性化し,IRF3/7のリン酸化を促進することによりIFN応答を誘導する.一方,三つ目のRLRファミリー分子であるlaboratory of genetics and physiology 2(LGP2)はCARDを欠如しており,単独ではRLR経路を活性化することはできない.にもかかわらず,LGP2はMDA5による二本鎖RNA認識を促進する働きがあることが示されている5–7).
このようにウイルス由来のRNAはTLRやRLRにより非自己として認識されるが,内因性RNAもまた免疫監視の対象となる.RNAのライフサイクルにおいて,内因性RNAは宿主因子による修飾や編集を受けることにより,自己RNAとしての特徴を獲得し免疫監視を免れている.しかし,非自己として誤認識された場合,過剰な免疫応答を引き起こし生体に悪影響を及ぼす.近年,RNA修飾に関する研究の目覚ましい進歩により,自己と非自己の識別に関する知見が蓄積しており,自己RNAに対する誤認識とそれによるI型IFNを中心とした慢性炎症との関連性が明らかになりつつある1, 8, 9).
本稿では,RNAを介した自然免疫応答の分子機構を,自己と非自己の識別における基本原理に言及しながら,宿主核酸認識機構と抗ウイルス応答について概説し,近年明らかになりつつある核酸修飾と免疫応答との関連と,核酸を介した免疫制御の開発について最近の我々の知見を含めて紹介する.DNA認識機構に関しては紙面の都合上,他誌を参考にされたい10–12).
RNAを認識するTLRとしてTLR3, TLR7, TLR8, TLR13が知られている.TLR3は合成二本鎖RNAアナログであるpolyinosinic-polycytidylic acid(poly I:C)を認識するRNAセンサーとしてよく研究されている.TLR3に認識されるpoly I:Cは少なくとも35 bp以上の長さが必要とされ,配列には依存しない13, 14).また,より長い二本鎖RNA(>90 bp)ではTLR3との結合が多価効果により安定化するとともに,TLR3分子間で多重複合体を形成することにより,効率的に下流のシグナルを活性化する15, 16).TLR3はマクロファージや樹状細胞といった免疫細胞以外にも,線維芽細胞,腸管上皮細胞,あるいは神経細胞を含むさまざまな非免疫細胞にも発現しており,主にウイルス感染時に生成される二本鎖RNAを認識し抗ウイルス応答を誘導する.TLR3欠損マウスは,ポリオウイルス,herpes simplex virus-1(HSV-1)やmurine cytomegalovirus(MCMV)といったウイルスに対して高い感受性を示すことが知られている17–19).また,ヒトTLR3の低機能型変異は,HSV-1によるヘルペス脳炎や重症インフルエンザ肺炎の罹患率と高い相関が確認されている20, 21).
TLR3と対照的に,TLR7やTLR8は一本鎖RNAを認識し,そのRNA結合様式や下流のシグナル経路も異なる.特に,イミダゾキノリン誘導体(イミキモドやレシキモド)はTLR7とTLR8のリガンドとしてよく知られている22).また,TLR7/8はポリウリジンやグアノシン・ウリジンに豊富なGU-rich配列によって活性化される23, 24).このような機能的な類似性に加え,TLR7とTLR8との間には構造的な類似点も認められている.TLR7とTLR8はともに,細胞外部ドメインに二つの特徴的なリガンド結合部位を有しており,TLR7は遊離グアノシン分子と一本鎖RNAに結合する一方,TLR8は遊離ウリジン分子と一本鎖RNAに高い親和性を示すことが,結晶構造解析により明らかにされている25–27)(図1).また,このようなTLR8の活性化にはエンドソームに局在するRNase T2やRNase 2によるRNA分解が重要である28, 29).同様に,TLR7に関しても,RNase T2に加え,5′エキソヌクレアーゼであるphospholipase(PLD)3/4によるグアニジンの効率的な生成が報告されている30).さらに,非免疫細胞にも発現が認められるTLR3と異なり,TLR7とTLR8は発現する細胞に明らかな違いが認められる31).TLR7はプラズマ様樹状細胞(plasmacytoid dendritic cell:pDC)やB細胞において高発現を示す.TLR8はヒト単球,コンベンショナル樹状細胞(conventional dendritic cell:cDC),好中球などの骨髄球系細胞に発現が認められるが,マウスTLR8はRNA認識に重要な配列が欠如しており一本鎖RNAに対する応答は認められない.これに加え,エンドソームにおける前述のRNaseの発現の違いもまた,これらTLRの活性に影響していると考えられる1).免疫系細胞に発現を示すことから,TLR7/8は一本鎖RNAをゲノムに有するインフルエンザAウイルス,水疱性口内炎ウイルス(vesicular stomatitis virus:VSV),severe acute respiratory syndrome coronavirus 2(SARS-CoV-2)などのウイルス感染に対してきわめて重要である32–35).特に,pDCはウイルス感染に対して,MyD88-TRAF6-IRF7を介して大量のI型IFNを産生する36, 37).ヒトにおけるTLR7欠損は血液中のpDCによるIFN応答が乏しく,COVID-19肺炎の重症化リスクと相関する38, 39).このようなウイルス感染に対する宿主防御的な機能にもかかわらず,TLR7活性化により産生されたI型IFNが肺組織中のマクロファージを刺激し,サイトカインストームを介して重症化を招くことも示唆されている35).それゆえ,TLR7/8の活性化の程度は感染症のコントロールにとってきわめて重要である.最近,TLR7が内因性リガンドとしてU11 small nuclear RNA(U11 snRNA)を認識することが報告されている40).興味深いことに,U11 snRNAはリボース2′-O-メチル化が欠如しており,この修飾を付加することでTLR7の活性化が抑制される.しかしながら,TLR7活性化のリスクがあるにもかかわらず,なぜU11 snRNAはリボース2′-O-メチル化が欠如しているのか明らかではない.U11 snRNAによるユニークなスプライシング機構が想定されているが,今後さらなる研究が必要である.
これらTLRに加えて,マウスに特異的なTLR13は細菌由来のリボソームRNA(rRNA)を認識し,感染防御に寄与している.TLR13は23S rRNAの特徴的な配列(CGGAAAGACCC, ACGGAAAGACCCC)を認識することに加え41, 42),このリガンド認識にはRNAステムループ構造が重要であることが,TLR13の構造解析により示されている43).またヒトTLR10は二本鎖RNAに加え,インフルエンザウイルスやhuman immunodeficiency virus(HIV)由来の構成成分を認識することが示唆されているが44–47),その生理的意義は明らかになっていない.
細胞質内のRNAは宿主細胞に対する脅威として捉えられ,RIG-IやMDA5といった二本鎖RNA監視機構により感知される.特にウイルス感染細胞において,ウイルス複製の中間体として生成される二本鎖RNAは,ウイルス感染の兆候としてこれらRNAセンサーに感知される.RIG-Iは300 bp以下の比較的短い二本鎖RNAを認識する一方,MDA5はより長い二本鎖RNAを認識する.このようなリガンドの長さに関する認識の違いは,それぞれが感知するウイルスの種類や認識の違いにも反映される.(+)鎖一本鎖RNAウイルスは,複製過程において大量の二本鎖RNAを生成することから,MDA5により感知される48, 49).一方,(−)鎖一本鎖RNAウイルスであるインフルエンザウイルスは,ゲノム内に生じるpanhandle構造と呼ばれる短い二本鎖領域,あるいは短い転写産物が二本鎖構造を形成することでRIG-Iに対するリガンドとなりうる50–53).また,レオウイルス科のウイルスゲノムは二本鎖RNAで構成されており,MDA5とRIG-Iの両方により感知される54).長さによる識別以外にも,5′末端におけるRNA修飾パターンはRIG-Iによる認識に重要である.典型的なRIG-Iのリガンドの特徴は,①5′末端における三リン酸(5′-PPP)あるいは二リン酸(5′-PP),②cap1, cap2における2′-O-メチル化の欠如,③平滑末端,として理解されている4)(図1).また,弱い相互作用ではあるが,5′末端の水酸基(5′-OH)に結合を示す一方,5′末端の一リン酸(5′-P)は感知しない.C末端ドメインは,5′PPP/PPとの結合部位を有し,2′-O-メチル化が欠如したN1との結合にも関与する.5′-OHと5′-Pとの間における反応性の違いは,RNaseによる分解産物の識別に重要である.tRNAやrRNAはRNaseによるプロセッシングを受け5′-P RNAを生成するが,このような自己RNAはRIG-Iによる認識を受けない.一方で,ribonuclease L(RNase L)によって分解されたウイルスRNAは5′-OHを生じるため,RIG-Iによる認識が可能である55).さらに,tRNA, rRNAに加えてDicerによる分解産物は,5′側あるいは3′側にオーバーハング配列を持つため,RIG-Iによる認識を回避できる.このようにRIG-Iの二本鎖RNA認識に必要な最短長は18~19 bpと非常に短いが,5′末端構造を認識することで感知可能となる.一方,MDA5によるRNA認識に関しては,末端構造の特徴は明らかになっておらず,長い二本鎖RNAであることが最も重要であると考えられている.
一方,このようなRNA免疫監視機構を回避するために,多くのウイルスが非自己RNAとしての特徴を無効化する巧妙な戦略を用いている.ハンタウイルスやブニヤウイルスを含む(−)一本鎖RNAウイルスは5′末端構造を5′-Pに変化させる一方で,アレナウイルスは5′末端構造にオーバーハング配列を付加することでRIG-Iによる認識を回避している56).さらに,インフルエンザウイルスを含むオルソミクソウイルスは,宿主RNAからキャップ構造を切り取り,自身のmRNAに取りつけるといったcap-snatchingと呼ばれる戦略を取り入れている57, 58).対照的に,コロナウイルス,ポックスウイルス,あるいはフラビウイルスなどのウイルスは,自身の2′-O-メチル化転移酵素によってN1メチル化(cap1)を誘導し,宿主RNAの5′末端構造を模倣する59).最近,N1メチル化を制御する宿主メチル化転移酵素であるmethyltransferase ribozyme 1(MTr1)を阻害することで,cap-snatchingウイルスの複製を阻害できることが報告されている60).
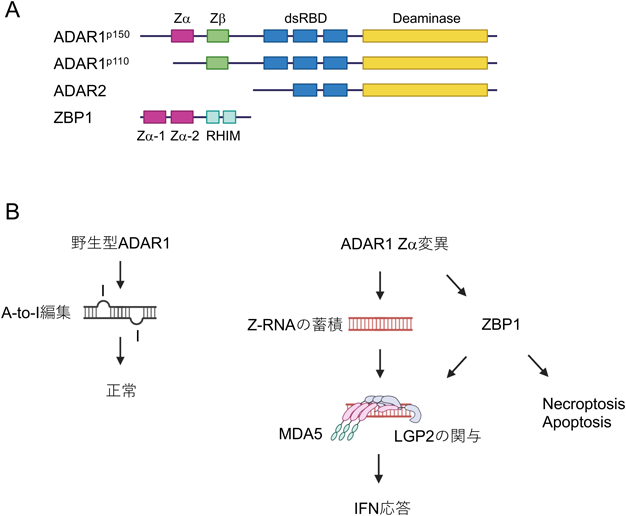
宿主ゲノムはLINE(long interspersed nuclear elements)やSINE(short interspersed nuclear elements)に分類される内因性レトロエレメントが全ゲノムの約50%を占める.代表的な要素として,LINE-1は約6000塩基の配列に逆転写酵素やエンドヌクレアーゼをコードしており,自ら複製してゲノム不安定を引き起こす可能性がある.一方,ヒトSINEにはAlu配列と呼ばれる約300塩基の配列が数多く散在しており,遺伝子発現に対して影響を与えることがある.特に,Alu配列が逆向きに配置されるとinverted Alu配列と呼ばれ,その転写物は潜在的に長い二本鎖RNAを形成しうることが知られている.基本的に正常な細胞では二本鎖RNAはほとんど検出されないが,何らかの原因により二本鎖RNAが細胞内に異常に蓄積すると,過剰なIFN応答を引き起こす61).このような病態は総称してI型インターフェロノパチーと呼ばれ,中でも単一遺伝疾患として希少ではあるが,Aicardi-Goutières syndrome(AGS)やSingleton-Merten syndromeが知られている8).AGSは重度の脳症と過剰な炎症応答を特徴とする先天性疾患であり,原因遺伝子によりさまざまな症状を呈する.現在,原因遺伝子として七つが報告されているが,特に近年ADAR1やIFIH1(MDA5)に関して病態形成に関わるメカニズムが明らかになってきた.adenosine deaminase acting on RNA1(ADAR1)は,アデノシン6位のアミノ基を脱アミノ化によりイノシンに変換するRNA編集(A-to-I編集)酵素である.イノシンはグアノシンとして認識されるため,宿主mRNAやウイルスRNAのコードを編集するほか,A-to-I編集はRNA二次構造を変化させることで,二本鎖RNA形成の抑制に重要であると考えられている62).またADAR1はp110とp150の二つのアイソフォームが存在し,それぞれ異なるプロモーターにより発現が制御されている(図2A).p110は核タンパク質として恒常的に発現しているが,p150は主に細胞質に局在を示し,IFN応答性に発現誘導が認められる.これらアイソフォームのうち,p150が二本鎖RNA形成の抑制に重要である.実際に,Adar1を欠損,あるいはp150のみを欠損した場合,強いI型IFN応答を特徴とするAGS様の病態を引き起こす63, 64).興味深いことに,このIFN応答はMDA5やMAVSの遺伝的欠損により正常化することが,二重欠損マウスの解析により明らかとなっている65–67).一方で,p110–/–単独欠損,あるいはp110–/–/Adar2–/–二重欠損ではISGの発現上昇は認められない68).にもかかわらず,これらのマウスは著しい致死性を示すことが明らかとなっており,特にイントロン内におけるRNA編集はp110とADAR2の欠如により著しく障害されることが確認されている68–70).さらに,p110/Adar2二重欠損の解析から,p150のA-to-I編集活性依存的なRNA編集が特に3′非翻訳領域(3′-UTR)内で同定されており,MDA5による認識を抑制していると考えられている68).
p150は,二本鎖RNA結合ドメイン,脱アミノ化ドメインに加え,N末端に細胞質移行シグナルを含むZ型RNA結合ドメイン(Zα)を有しているが,p110には存在しない(図2A).ADAR1のZαドメインはCGリピートで構成される左巻きZ-RNAに対して結合能を示し,p150によるA-to-I編集活性に影響を与えると考えられている.最近p150における生物学的意義に関して,Zαドメインにおける変異マウスを用いた解析により,その重要性が明らかになってきた(図2B).P195A変異(ヒトP193A)のホモ接合体は表現型を示さないが,ADAR1あるいはp150のヘテロ接合体に対してP195A変異が挿入されると,マウスは肝臓,腎臓,脾臓などでIFN応答を呈し致死性を示す71).興味深いことに,この表現型はMDA5欠損のみならず,LGP2欠損によっても改善する.LGP2はMDA5の重合形成に寄与することから,MDA5による自己RNAに対する認識への関与が示唆される.一方でTangらは,種間で保存されたN175とY179に変異を導入したマウスを作製したところ,特に肺などにIGSの発現増加が認められたが,AGSに典型的な病態は示さず致死性も示さないことを報告している72).またこのようなIFNシグナルの活性化はインフルエンザウイルス感染に対して抵抗性を示すことを明らかにしている.さらに中濱らは,W197A変異マウスを作製し,AGSにみられる脳症を呈していること,特に脳組織においてISGの強い発現誘導が認められ,Adar1P195A/nullと同様に致死性を示すが,MDA5欠損により改善できることを示した73).また脳組織では,ある特定の3′-UTRにZαドメインが関与するA-to-I編集部位の存在が示唆されているが,その全貌は明らかになっていない.
ADAR1以外にも,左巻き核酸を認識するタンパク質としてZ-DNA binding protein 1(ZBP1)が知られている(図2A).ZBP1のZαドメインはDNAウイルスによって生成される左巻きZ-DNAを認識すると考えられていたが,ノックアウトマウスの解析からDNAに対するIFN応答には関与しないことが明らかになっている74).むしろZBP1は,インフルエンザなどのウイルス感染に対して,receptor-interacting protein kinase 3(RIPK3)と会合しmixed-lineage kinase-like(MLKL)のリン酸化を介して細胞死の形態の一つであるnecroptosisを誘導する,あるいはRIPK1を活性化しCaspase-8を介したapoptosisの誘導を制御する75, 76).またZBP1はNLRP3依存的なインフラマソームを活性化することでpyroptosisと呼ばれる細胞死にも関与する77).一方で,ADAR1との機能的な類似性からADAR1 Zα変異により引き起こされるIFN応答への関与が想定されていたが,注目すべきことに,ADAR1 Zαドメイン変異(Adar1P195A/p150null)によって誘導される致死性がZBP1の遺伝的欠如(Zbp1–/–)によって著しく改善することが報告されている.Adar1P195A/p150nullマウスで観察されるISG発現増加に関しても,Mavs–/–と比べて部分的ではあるが,Zbp1–/–により緩和されることから,MDA5-MAVS依存的なIFN応答の活性化への関与が示唆される.一方で,3′-5′ DNAエキソヌクレアーゼであるTREX1の欠損は,細胞質に異常に蓄積したDNAが著しいI型IFNを誘導することが知られているが,ZBP1はDNA依存的に形成された慢性炎症の病態には関与しないことが示されている78).また,ZBP1のMDA5-MAVS非依存的な関与も示唆されている.Adar1–/–Mda5–/–やAdar1–/–Mavs–/–マウスはAdar1–/–による致死性を改善するものの,依然新生仔死亡となるが,Adar1–/–Zbp1–/–Mavs–/–三重欠損マウスでは40%が生存可能である79).この背景にあるメカニズムとして,ZBP1によるRIPK3依存的な細胞死の関与が指摘されている.マクロファージにおいてZBP1はIFN依存的にADAR1と会合するが,RIPK3との相互作用は認められない80).しかし,ADAR1の細胞質移行を阻害した場合,RIPK3はZBP1と相互作用すること,さらにRIPK3を欠損するとZBP1とADAR1の相互作用が促進し細胞死が抑制されることから,Adar1欠損マウスではZBP1がRIPK3を介して細胞死を誘導することが示唆される80).実際に,Ripk3–/–Adar1P195A/p150nullマウスは,Adar1P195A/p150nullマウスと比較し生存率が改善することから,RIPK3はZBP1の機能の少なくとも一部を説明すると考えられる81).一方で,ADAR1とZBP1は,ともにZ-RNAに対して結合能を示すことから,細胞内ではZ-RNAに対して競合的に認識している可能性がある64).つまり,ADAR1はZ-RNAと結合しZ-RNAの蓄積を抑制する一方で,ZBP1はZ-RNAの認識を介してRIPK3依存的な細胞死を制御している.しかしながら,in vivoにおいてZBP1がどのように細胞死やIFN応答を制御しているかに関しては明確に証明されておらず,より詳細な分子機構の解明が必要である.
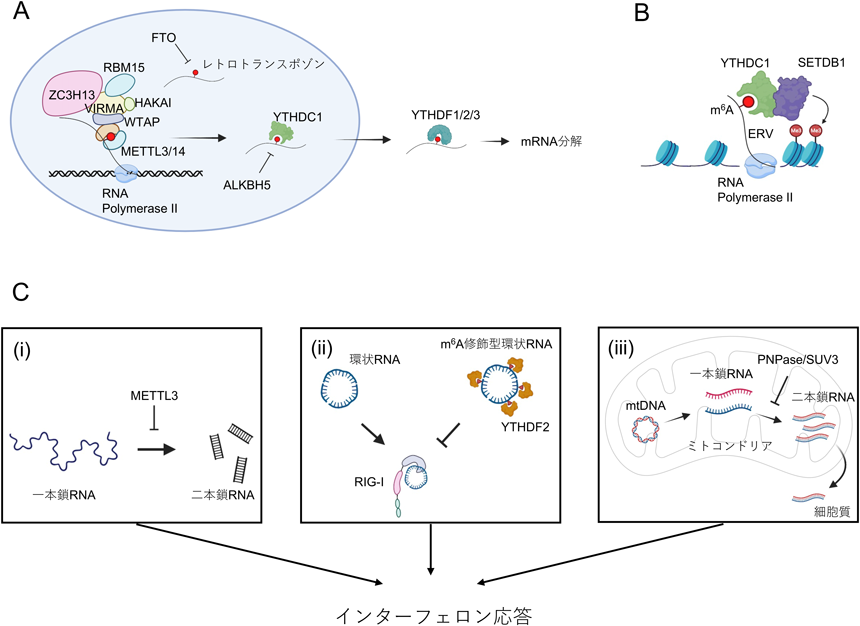
RNAは細胞内で150種類にも及ぶ化学修飾を受けており,遺伝子発現制御において重要な役割を果たしている82).中でもN6-methyladenosine(m6A)修飾は,細胞内で高頻度に認められるRNA修飾であり,ポリA鎖が付加されたRNAに含まれるアデノシン全体の0.2~0.6%に存在する83).細胞内では複数のm6Aメチル基転移酵素が存在し,それぞれ基質となるRNAや標的配列,その生物学的意義が異なる.特にmRNA上のm6A修飾は,メチル基転移酵素であるMETTL3によって付加される.METTL3は細胞内でMETTL14とヘテロ二量体を形成し,さらにWTAP, VIRMA, HAKAI, RBM15, ZC3H13といったアダプタータンパク質から構成されるm6A修飾酵素複合体(writer complex)として機能する(図3).また,METTL3によるm6A修飾は,核内で転写とほぼ同時に起こり,コンセンサス配列DRACH(D=A/G/U, R=プリン,A=m6A, H=A/C/U)のアデノシンに対してメチル基が付加される9).この修飾は主に,mRNAの終止コドン付近や3′-UTRに高頻度に認められるが,すべてのDRACH配列が修飾を受けるわけではなく(~20%)9),またその頻度も一定ではない.m6A修飾は一群のreaderタンパク質により読み取られ,その機能によりRNA代謝に与える影響はさまざまである9, 84)(図3).特にYTHドメインを持つreaderタンパク質の機能はよく研究されている.中でもYTHDF1, YTHDF2, YTHDF3は細胞質でm6Aを認識するreaderタンパク質である.YTHDF1は翻訳開始因子eIF3との会合を介して翻訳を促進する一方,YTHDF2はmRNA分解を促進する85, 86).YTHDF3はYTHDF1と協調的に翻訳促進に寄与すると考えられている87).しかしながら,YTHドメインによるm6A結合は構造学的にきわめて類似していることから,このようなYTHファミリー間における機能的な相違に関して疑問が生じている.最近の研究により,YTHDF1/2/3はm6A認識部位に関して明らかな差異はなく機能的に重複しており,全体としてmRNA分解を促進するというモデルが提唱されている88–90).一方,核内のm6A認識にはYTHDC1が主に関与しており,スプライシング,核–細胞質輸送,RNA分解,エピジェネティック制御など,その機能は多岐にわたる91–93).加えて,ALKBH5やfat mass and obesity-associated protein(FTO)といったm6A脱メチル化酵素(eraser)によりm6A修飾は可逆的に制御される.さらに,m6A修飾はRNA二次構造を緩和する方向に働き,hnRNPC, hnRNPA2B1, IGF2BP3といったRNA結合タンパク質によるRNA認識に影響を与えている可能性が指摘されており94–96),m6Aを介した遺伝子発現制御の複雑性が示唆される.本節では,自然免疫応答,特に抗ウイルス応答との関連に加え,自己と非自己の識別におけるm6Aの役割に関して最新の知見を述べる.
1)m6A修飾によるIFNシグナル制御
m6A修飾が免疫応答に及ぼす影響は,抗ウイルス応答においてよく研究されている.m6A writer complexは,IFNやISGをコードするmRNAに対してm6Aを付加し,抗ウイルス応答を調節している.具体的に,ウイルス感染細胞ではIFNBやIFNAなどのmRNAはm6A修飾を受けることが報告されている97, 98).METTL3欠損細胞では,これらmRNAの安定化を介してIFN応答が増強することで,結果としてウイルス複製,および伝播が抑制される.逆に,m6AをmRNAから除去するいわゆるeraserとして機能するタンパク質の一つであるALKBH5を欠損した細胞では,IFNB mRNAの発現が低下しウイルス感染が悪化する97).このように,m6A修飾はIFN mRNAを標的とし不安定化を促進することで,IFN応答を制限する役割を担っている.また,一方でm6Aは抗ウイルス免疫を促進することも報告されている.ウイルス感染細胞では,MAVS, TRAF3, TRAF6をコードするmRNAがDDX46により認識され,そのRNAにALBKH5が結合,mRNAに付加されたm6Aを脱メチル化する.この結果,これらmRNAの核から細胞質への輸送が阻害され,I型IFN産生が抑制される99).さらに,ウイルスDNAに対する核内認識タンパク質として同定されたhnRNPA2B1は,別のm6A eraserであるFTOと恒常的に結合しているが,ウイルス感染が起こると解離しCGAS, IFI16, STINGのmRNAに付加されたm6Aを保護することで,核–細胞質輸送を促進し,抗ウイルス応答を高める100).これらの知見は,m6A修飾がmRNAの核–細胞質輸送を制御して抗ウイルス応答に重要な役割を果たしていることを意味する.しかし,ALKBH5とFTOの機能的な違いについては未解明な部分が多く,さらなる研究が必要である.
ALKBH5によるmRNA安定性制御を介した抗ウイルス応答の調節機構も報告されている.ウイルス感染時には,ALKBH5のアルギニン残基(R107)の脱メチル化によって脱メチル化酵素活性が低下し,α-ケトグルタル酸デヒドロゲナーゼOGDH mRNAにおけるm6A修飾が増加し,mRNA分解が促進する.この変化が最終的に抗ウイルス代謝環境を誘導しウイルス複製を抑制するとされている101).しかしながら,この結果は前述の抗ウイルス応答におけるALKBH5の役割との矛盾が指摘されている.ALKBH5が標的遺伝子の脱メチル化を促進する一方で,酵素活性の低下によるm6A修飾の増加がいかに両立するのかは不明である.さらに,これらのメカニズムのうち,どちらが抗ウイルス応答に重要な役割を果たしているのかについては,今後のさらなる研究が必要と考えられる.
2)ウイルスRNAに対するm6A修飾
細胞内に侵入したウイルスRNAもまた,宿主RNAと同様にm6A修飾を受ける.核内で複製するウイルスの多くは,自身のRNAの転写が起こる際に,宿主m6A writer complexを利用すると考えられる.例外として,B型肝炎ウイルス(HBV)におけるm6A修飾には,ウイルス由来HB Xタンパク質によるMETTL3/14との会合が必要である102).一方,細胞質で複製するウイルスの場合,核内に局在するm6A writer complexがいかに細胞質へ移行するか,不明な点が多い.C型肝炎ウイルス(HCV),VSV, SARS-CoV-2では,実際にm6A writer complexの細胞質への局在変化が確認されている103–105).これにはRNA依存的RNAポリメラーゼの一時的な核内移行の関与が想定されているが106),現在のところm6A writer complexの細胞質移行に関するメカニズムは明らかにされていない.
このようなウイルスRNAにおけるm6A修飾は,宿主RNAを模倣することで細胞内RNAセンサーの監視機構を逃避する上で重要であると考えられる.たとえば,VSVウイルスRNAに対するm6A修飾は二本鎖RNA形成を緩和させる103).HBVやHCVのRNAに対してm6A修飾は,YTHDF2やYTHDF3といったm6A readerタンパク質との相互作用により,RIG-Iによる認識を回避する107).ほかにも,RSウイルス,SendaiウイルスやSARS-CoV-2ウイルスなどさまざまなウイルスがm6A修飾を利用した免疫回避戦略を利用している108–110).しかし,いかにウイルスRNAがm6A修飾を用いて宿主RNA監視機構から逃避しているか,詳細なメカニズムはよくわかっていない.
3)m6A修飾を介した内因性RNAによる自然免疫応答
mRNAワクチンに応用されているN1-methyl-pseudo-uridine(N1mψ)の重要性は広く認知されるようになったが,m6A修飾もまた免疫回避にとって重要な核酸修飾の一つである.たとえば,内因性レトロウイルス(endogenous retrovirus:ERV)由来の転写産物はm6A修飾の重要な標的であり,m6A修飾によりERVの発現量が調節されている.Mettl3やMettl14を欠如した胚性幹細胞(embryonic stem cell:ESC)は,マウスERVの一種であるintracisternal A-particles(IAP)mRNAが安定化し,その結果発現が増加することが示されている111).また,ESCにおいてFTOはLINE1のm6Aを脱メチル化することが報告されている93).Fto欠損ESCでは,メチル化されたLINE1由来の転写物がYTHDC1により認識され,分解されるか,あるいはSETDB1によるクロマチンリモデリングを介してERVの発現が抑制される(図3A, B).このことから,m6A修飾はERVの細胞内蓄積を防ぐ上で重要な役割を果たしており,哺乳類の胚発生において何らかの意義が示唆される.さらに,血球系細胞特異的にMettl3を欠損させたマウスでは,胎仔肝臓における顕著な造血障害のため胎生後期より致死となる112).興味深いことに,Mettl3欠損造血幹細胞の細胞内には二本鎖RNAの蓄積が観察され過剰なIFN応答が認められることから,m6A修飾は二本鎖RNA形成を抑制し,胎生期の造血を維持していると考えられる112)(図3C).環状RNAにおけるm6A修飾もまた,自己と非自己の認識に関与している.環状RNAは,pre-mRNAが成熟する過程において,通常とは逆方向のスプライシング(back splicing)により生成され,全mRNA発現量の2~4%存在すると考えられている113–116).興味深いことに,in vitroで転写された環状RNAを細胞内に導入すると,RIG-Iを介したIFN応答が誘導されるが,最近の研究により,その活性化抑制には環状RNA上のm6A修飾が関与していることが報告されている117, 118).m6Aが付加された環状RNAは,readerタンパク質であるYTHDF2が直接結合することで,RIG-Iを介した免疫応答を抑制することが可能である117)(図3C).しかし,RIG-Iはm6A修飾型環状RNAを認識するものの,MAVS活性化にはむしろRIG-Iの構造変化が重要であることが示唆されている.また,このようなRIG-Iの構造変化にはK63型ポリユビキチン鎖の存在が必要であるが,なぜ一本鎖RNAである環状RNAがRIG-Iに認識されるのか,その詳細なメカニズムは解明されていない.
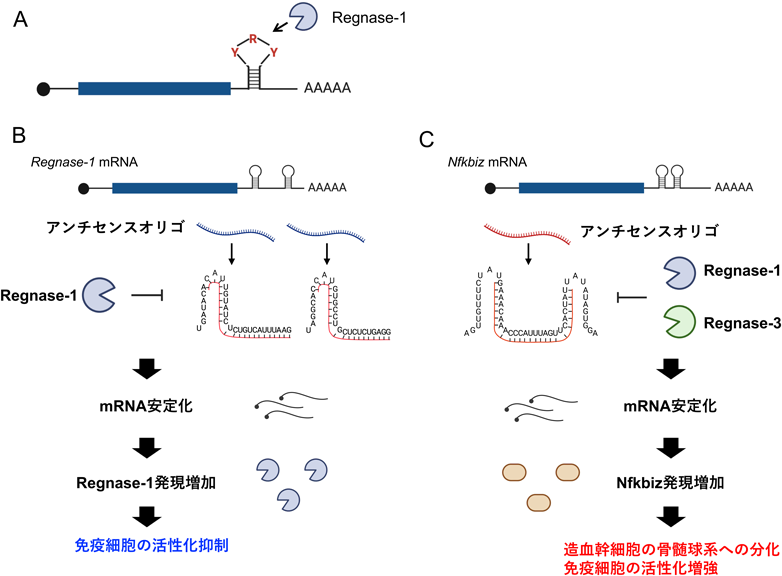
疾患制御を目的としたオリゴ核酸は一般に核酸医薬として近年注目が高まっている.そのほとんどは,疾患原因遺伝子に対するmRNA発現抑制やエクソンスキッピングを目的としたアンチセンスオリゴ核酸である.従来TLRやRLRを標的とした免疫制御に関する報告は数多く,現在も開発が進められているが,筆者らは独自のmRNA分解機構に着目し,オリゴ核酸を用いた免疫制御法を開発してきた(図4).mRNA分解酵素であるRegnase-1は,炎症性サイトカインや免疫細胞活性化に関与するタンパク質をコードするmRNAを標的として分解し,免疫細胞の活性化に対してブレーキとして働く122).Regnase-1を介したmRNA分解には,3′-UTRに存在するピリミジン–プリン–ピリミジン配列をループに持つステムループ構造の認識が必要である123).最近の筆者らの研究により,Regnase-1が標的とするステムループ構造を破壊するアンチセンスオリゴ核酸(ASO)を開発し,これが標的遺伝子のmRNA分解抑制を介してタンパク質発現増加を誘導可能であることを見いだした124).Regnase-1は自身のRegnase-1 mRNAにも2か所の標的ステムループ配列が存在しており,mRNA分解によるRegnase-1発現のオートフィードバック機構が存在することを明らかにしている.そこで,これら2か所の標的配列に対してASOを設計し細胞導入したところ,Regnase-1の自身による分解を抑制することでその発現を増加させ,さらにマクロファージによる炎症誘導を抑制できることを明らかにした.さらに,マウス個体において急性呼吸促迫症候群や,肺線維症,多発性硬化症モデルマウスの疾患制御も可能であることから,Regnase-1標的ASOは慢性炎症の病態制御に有用であることが示唆される124).一方で,Regnase-1 mRNA以外のRegnase-1標的遺伝子に対しても同様に,発現をコントロールすることが可能である.最近,Regnase-1とそのファミリー分子であるRegnase-3は共通のステムループを認識していることから,これら二つのRegnaseが造血の分化方向性を制御することを明らかにしている125).重要なことに,Nfkbiz mRNAはRegnase-1とRegnase-3の間における共通の標的遺伝子であり,RegnaseによるNfkbiz mRNA分解が造血幹細胞の分化方向性を決定づける新たな制御相であることを明らかにした.興味深いことに,Nfkbiz mRNAの3′-UTRにも標的ステムループが2か所存在し,これら二つのステムループが短いスペーサー配列(13 nt)を挟んで連続している.そこで,このステムループを破壊するASOを設計し造血幹細胞に導入したところ,骨髄球系細胞への分化バイアスが誘導されること,さらにIL-1βやTNFといった炎症応答に対して骨髄球系への分化を加速させることを明らかにした125).興味深いことに,このASOは造血幹細胞以外にも,マクロファージに対する炎症応答増強作用や,T細胞に対して強いエフェクター機能と増殖を誘導できることを見いだしており,今後抗腫瘍免疫増強,あるいはワクチンアジュバンドとしての効果が期待される.