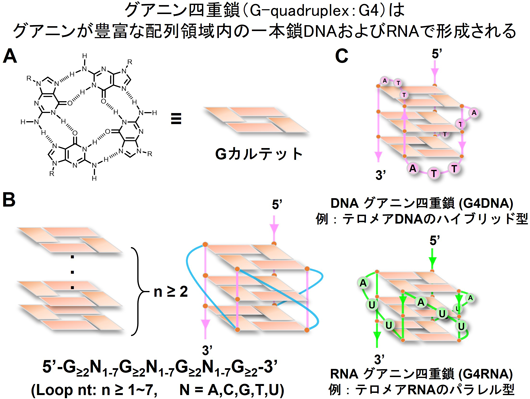
DNA・RNA高次構造には多様性がある.DNAは,右巻き二重らせんであることがワトソン博士とクリック博士によって1953年に報告された1).このDNAの基本的な構造は「B型DNA」と呼ばれている.一般的に知られているこの構造以外にも,左巻き(Z型)DNA,三重鎖(H型)DNAなど「非B型DNA」と呼ばれる構造が発見されており,DNAはその配列の特徴や溶媒の環境によりさまざまな構造を形成する2).非B型DNAの一つであるグアニン四重鎖(G-quadruplex:G4)は,グアニンが豊富な配列領域内の一本鎖DNAやRNAで形成される(本稿では各々G4DNA, G4RNAと略称)(図1).各グアニンが二つの隣接するグアニンと非ワトソン・クリック塩基対であるフーグスティーン塩基対を形成し,四つのグアニン分子が「Gカルテット」と呼ばれる正方形の平面配置をとる(図1A).さらにGカルテットは,互いの上に積み重なりG4を形成する.G4は,少なくとも二つの隣接グアニンを有する四つのトラクトと三つのループ領域で形成される(図1B)3).
G4の形成は,カチオンの結合・温度・配向・ループサイズなど,いくつかの要因により影響を受ける4).G4はGカルテットの中心に一価の陽イオンが結合することで安定化する.一般的には,K+>Na+>Li+の順にG4の安定化作用が強い5).ループ長および配列によりさまざまなG4が形成され,分子内構造,二分子構造,四分子構造,高次Gカルテット構造,ストランドが同一の配向を持つパラレル型,4本のうち1本だけ逆を向くハイブリッド型,配向が2本ずつ交互になるアンチパラレル型などが報告されている6–8).G4RNAはG4DNAより熱力学的に安定で構造的多様性が低い9).1~5ヌクレオチドのさまざまなループ長のRNAオリゴヌクレオチド・ライブラリーを使用した実験の結果,G4RNAトポロジーはループ長には無関係にパラレル型を形成することが示されている10).例として,テロメアの構造があげられる.テロメアは,TTAGGG配列の4リピートでK+結合によりハイブリッド型G4DNAを形成し,UUAGGG配列の4リピートでパラレル型G4RNAを形成する(図1C).重要な点として,上述のG4トポロジー解析はすべてin vitroによる結果であり,生物物理学的な解析結果がin vivoにおいても適応されるのか,そして生体内イベントにどのように関与するのかは未解明である.
これまでに,G4を解析するための実験技術が多数報告されている.それらは,in vitroにおける生物物理学的解析,in silicoによる予測解析,ゲノムDNA・RNAの網羅的解析(G4DNA・G4RNAプロファイリング)に大別される.これらの手法は一長一短であり,G4の形成を確認するにはいくつかの解析法を組み合わせることが重要である.
1)in vitroにおける生物物理学的解析
X線結晶構造解析法,核磁気共鳴分光法(nuclear magnetic resonance:NMR),円二色性分光法(circular dichroism:CD),可視・紫外(UV-Vis)吸収スペクトル融解法,核酸の切断を利用したフットプリント法などが用いられる.G4の詳細な分子構造を明らかにできる手法はX線結晶構造解析法であるが,結晶を作製するには大量の核酸が必要であることや,結晶形成のためのさまざまな添加物(特に塩)がG4の分子構造に影響を与えることから,解析条件により結果が異なることがある.たとえば,前節で述べたテロメアの構造解析も塩により結果が異なり,K+結合でハイブリッド型,Na+結合でアンチパラレル型を形成する11).NMRにおいても大量の核酸が必要であることや,解析できる塩基長に限界がある.G4解析に頻用される手法としてCDがある.CDの利点として,比較的少ない量の核酸で測定が可能であること,測定条件を簡易的に変えられること,短時間で測定可能であることがあげられる.CDは,X線構造解析のように詳細な分子構造を明らかにすることはできないが,G4のトポロジー(パラレル型,アンチパラレル型,ハイブリッド型)を容易に決定することができる.核酸の切断を利用したフットプリント法によりG4の構造を推定することもできる.G4DNAの解析にはジメチル硫酸(dimethyl sulfate:DMS)フットプリント法を用いる.DMSはグアニンのN7位をメチル化する.DNAにDMSを処置後,ピペリジンを添加することでメチル化部位での切断が誘導される.その際,Gカルテットを形成するグアニンはフーグスティーン塩基対を形成しているためN7位のDMSによるメチル化に対する抵抗性を有し,ピペリジンによる切断が阻害される.G4RNAの解析にはRNase T1フットプリント法を用いる.RNase T1は一本鎖RNAのグアニンに対して特異性を示すため,G4やヘアピン形成に関与するグアニンは抵抗性を有する.UV-Visスペクトルを用いた解析法では,スペクトルの温度変化からG4の融解曲線を構築し,構造安定性を反映する熱安定性Tm,エンタルピー変化量ΔH°,エントロピー変化量ΔS°,ギブス自由エネルギー変化量ΔG°の熱力学的パラメータを算出することができる.
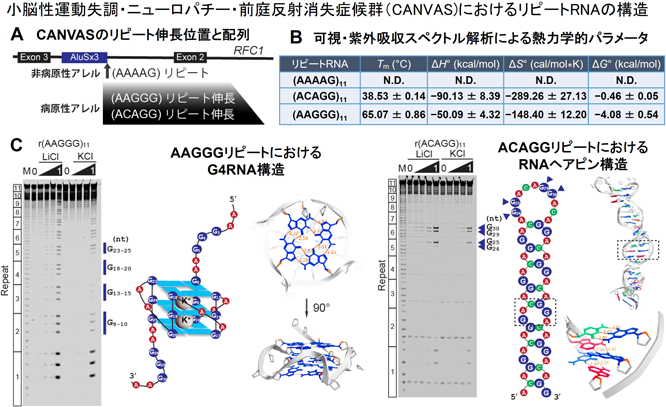
例として,筆者らが報告した生物物理学的解析を紹介する.小脳性運動失調・ニューロパチー・前庭反射消失症候群(CANVAS)は,replication factor C subunit 1(RFC1)のイントロン2に存在するAAAAGリピートがAAGGGもしくはACAGGリピートに変異し伸長することで発症する遺伝性の神経変性疾患である(図2A)12, 13).興味深いことに,伸長したリピートの「配列」によって病態の症状に違いがみられる.筆者らは,病原性リピートRNAを合成しin vitroにおける生物物理学的解析を行った.結果として,病原性リピートRNAは,非病原性リピートRNAと比較して「硬いRNA構造」を形成することが熱力学的パラメータで明らかになった(図2B).さらに,AAGGGリピートRNAではG4RNAの形成が,ACAGGリピートRNAではG–C塩基対結合に加えてG : A塩基対結合により安定化したミスマッチヘアピンRNAの形成が観察された(図2C)14).RFC1リピート伸長の構造多型に関するこれらの知見は,CANVASの病態メカニズムに関与する可能性がある.今後,CANVAS患者由来iPS神経細胞や疾患モデルマウスを用いてこれらRNA構造と神経機能異常との関連性を解析する必要がある.
2)in silicoによるG4予測解析
G4のin silico解析に関しては,Quad-Parser15),QGRS Mapper16),G4P Calculator17),Quadbase18),cGcC score19),G4Hunter20)など,これまでに多くの予測アルゴリズムが開発されている.これらは,G4形成の可能性を有する配列potential G-quadruplex-forming sequences(PQS)を同定できる.HeLa細胞,ヒト線維芽細胞,ヒトES細胞,およびヒトiPS細胞を用いたin silico予測の結果,PQSはヒトゲノムに約300,000か所存在し,プロモーター・CpGアイランド・複製起点・ヌクレオソームフリー領域・5′非翻訳領域(5′-untranslated region:5′-UTR)・第1エクソンの位置と高度に相関していることが示された15, 17, 20, 21).Quad-Parserを使用したin silico予測では,米国National Institutesof Health(NIH)に登録された既知の遺伝子38,915個の5′-UTRのうち,約1割にPQSが同定された15, 22).しかしながら,in silico予測はDNAの一次配列からG4コンセンサス配列「G2–3+N1–7 G2–3+N1–7 G2–3+N1–7 G2–3(N=A, C, G, T)」に基づき結果を導き出すため,バルジ型モチーフ(非グアニン塩基が三つのGトラクトの中に含まれるモチーフ)23)の予測には不向きである.さらに,in silico予測は3次元的にG4が形成されていることを確認できない.それらの問題を克服するため,G4コンセンサス配列に制限されることなくin silico予測ができるアルゴリズム「G4RNA screener」が開発された24, 25).G4RNA screenerは,cGcC score, G4Hunter, G4NNの三つからなるin silico予測法である.G4NNは,実験的なエビデンスを含めた機械学習を利用しており,人工知能は今後のin silicoによるG4予測解析の主流になると思われる.
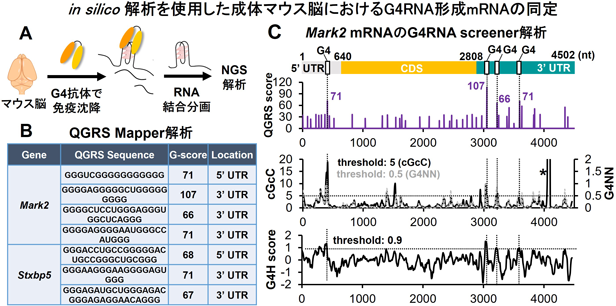
例として,G4のin silico解析を使用した筆者らの論文を紹介する.筆者らは,成体マウスの前脳組織におけるG4RNA形成mRNAの同定を試みた.G4特異的抗体を用いたRNA免疫沈降によりG4RNA形成mRNAを濃縮しシーケンス解析を行った(図3A).QGRS Mapperを適用したところ,同定されたすべてのmRNAは2層または3層のGカルテットからなるG4RNAを有することが予測された.1分子のmRNA中に複数のG4RNAを形成する可能性があるMark2とStxbp5に着目し(図3B),G4RNA screenerを適用した.QGRS Mapperで予測されたG4RNA形成配列の領域は,G4RNA screenerでも高いG4RNA形成スコアを示すことが確認された(図3C).これらの予測されたG4RNA形成配列は,前項で紹介したCDとRNase T1フットプリント法を用いることで,実際にG4RNAを形成することを確認した26).
3)G4DNAプロファイリング
ゲノムワイドにG4DNAを同定するためのアプローチとして,ポリメラーゼ・ストップアッセイが最初に報告された27).この手法は,G4結合性リガンドであるピリドスタチンもしくはK+の添加によりG4の構造を安定化させ,シーケンス時にG4DNAの位置でポリメラーゼの進行が停止することを利用する.ポリメラーゼ・ストップアッセイにより,ヒトBリンパ球細胞において約700,000か所のG4DNAが同定された.さらにその後,複数の生物種での解析も実施され,解析した12の生物種すべてでG4DNAの形成が確認された28).しかしながら,ポリメラーゼ・ストップアッセイは裸のDNAで解析するためクロマチン内のG4DNAを捉えていない.
クロマチン内のG4DNAを同定するために,G4特異的抗体BG429)を用いたG4ChIP-seqが開発された30).この手法により,ケラチノサイト株HaCaT細胞では約10,000か所のG4DNAが同定された.また,G4DNAピークはユークロマチンにおけるヌクレオソームフリー領域に集中しており,G4DNA形成と転写活性に正の相関関係がみられた30).一方,G4ChIP-seqはホルムアルデヒド固定や超音波処理などの実験条件により,G4DNAの形成に影響を与える可能性がある.また,実験には多数の細胞を必要とする.これらの課題を克服するため,生理的な条件下でG4DNAを捉えるための代替アプローチが開発されている.Cleavage Under Targets and Tagmentation(CUT&Tag)31)を応用したG4 CUT&Tag32, 33)は,非固定条件で細胞中のG4DNAをG4特異的抗体により捉えシーケンスが可能である.G4 CUT&Tagは,G4 ChIP-seqよりも分解能が向上しピーク数が約2倍に増加した.さらに,G4 CUT&Tagを単一細胞に適用したsingle-nuclei G4 CUT&Tag34)も報告されているが,2種類の培養細胞株を分離することにとどまっており,複雑な組織ではいまだ実施されていない.
細胞内のG4DNAおよびG4RNAを検出するためにG4特異的抗体を開発することは,CUT&TagによるG4DNAゲノムマッピングだけでなく,細胞内イメージングや免疫組織染色にも適用できることから有益であると思われる.一方で,これまで報告された数種のG4特異的抗体は,すべて特定のG4コンホメーションに対して結合優先性を示す35).たとえば,頻用されるG4特異的抗体BG4はパラレル型に対して高い親和性を持つ.また,BG4は大腸菌から精製する単鎖可変領域フラグメント(single-chain variable fragment:scFv)であるが,精製の難易度が高くロット間でG4とのアフィニティのばらつきが大きい.G4特異的抗体の代用としてG4結合性リガンド(ピリドスタチンやPhenDC3)にビオチンを有機化学合成により付加したプローブによるCUT&Tag「Chem-map」がある36).Chem-mapもG4DNAを検出できるが,G4結合性リガンドにはG4DNAと二本鎖DNAの動的平衡を解消し,G4DNAを人為的に形成させる性質がある.また,Chem-mapのプローブ合成には有機化学の知識と設備が必要である.その他,G4DNAが細胞内のオープンクロマチンに多く形成される性質を利用したヌクレアーゼ法(S1ヌクレアーゼ37),マイクロコッカル・ヌクレアーゼ38))がある.しかしながら,これらヌクレアーゼ処理は非生理的条件下で行われることや,得られる配列がオープンクロマチンに限定されること,G4特異性がないことが問題としてあげられる.
上述のとおり,細胞内G4DNAプロファイリングは,G4特異的抗体・G4結合性リガンド・ヌクレアーゼを使用する手法が開発されている.しかしながら,これらの結果を解釈する際には依然として慎重な評価が必要であり,より正確で簡便なG4DNAゲノムマッピング法が求められている.
4)G4RNAプロファイリング
in vitroにおけるG4RNAプロファイリングとして,逆転写(reverse transcription:RT)の停止を利用したシーケンス法rG4-seq39)とRT停止プロファイリング40)が報告された.手法の詳細は割愛するが,HeLa細胞におけるrG4-seqの結果では,mRNAで合計3383か所のG4RNAが同定された.そのうち3′ UTRには2086か所みられ,5′ UTR(540か所)やコーディング領域(697か所)と比較して多く観察された.RT停止プロファイリングでは,HEK293T細胞およびHeLa細胞でそれぞれ12,009か所および12,035か所のG4RNA(それぞれ6506か所および6281か所は非重複)を同定した.各領域の合計長で正規化すると,G4RNAの密度はコーディング領域よりもUTRで4~5倍程度高かった.また,G4RNAはmicroRNA(miRNA)標的サイトとポリアデニル化サイトの近くに豊富に局在したことから,miRNAを介した制御と代替ポリアデニル化におけるG4RNAの役割が示唆された.
細胞内G4RNAプロファイリングには二つの手法が提案されているが40),いずれもin vitro RT停止プロファイリングを応用している.つまり,細胞内でグアニンを何らかの方法で修飾し,その後in vitro RT停止プロファイリングを実施する.一つ目は,リボースの2′-ヒドロキシル基を修飾する試薬(selective 2′-hydroxyl acylation analyzed by primer extension:SHAPE)を用いた解析法である.SHAPE試薬の一つである2-methylnicotinic acid imidazolide(NAI)とRNAを細胞内で反応させる.G4RNAではGトラクトの最後のグアニンの2′-ヒドロキシ基が露出することから,細胞内でNAIにより優先的にアシル化される.その後,in vitro RT停止プロファイリングにより修飾グアニンを同定する.二つ目は,細胞をDMSで処理しグアニンのN7位をメチル化する手法である.細胞内で形成されたG4RNAは,Gカルテット形成に関与するグアニンがDMSのメチル化に対して抵抗性を有する.その後,抽出されたRNAに対してK+条件下でG4RNAをリフォールディングさせ,RT停止プロファイリングを行う.細胞内でG4RNAが非形成の場合,グアニンがメチル化されるため,in vitroのK+条件下でもG4RNAのリフォールディングは起こらない.
これら二つの手法を用いてマウス,ヒト,酵母を解析したところ,ほとんどのG4RNAが細胞内で形成されていないことが報告された40).しかしながら,これらの手法は試薬反応時間枠内のRNA構造立体配座を分析するため,個々のG4RNAの構造立体配座や動的な構造相互変換などを反映できていないことから結果が疑問視されている41).G4RNAの動的構造を観察するには時間分解能を向上させたアプローチが必要になる.
1)G4DNAの生物学的機能
G4DNAは,テロメアの維持・DNA複製・ゲノム不安定性・転写との関連性が報告されている.テロメアがG4DNAを形成することは,G4DNAがテロメラーゼを介したテロメアの伸長と関連する可能性を示している42).テロメアG4DNAはテロメラーゼの機能を阻害すると考えられていたが43),DNAヘリカーゼであるRTEL1はテロメアG4DNAを解消することによってテロメアを維持すること44),また,G4DNAがテロメラーゼの足場として機能することも報告されている45).テロメアの維持を阻害することでがん細胞の増殖抑制効果を狙ったG4結合性リガンドの探索が行われているが,テロメアにおけるG4DNAの詳細な生理機能は未解明な点が多い.
G4DNAはゲノム不安定性にも関与する.DNAヘリカーゼであるPIF1は,ゲノム不安定性によるG4DNAを介したDNA切断を阻害する.さらに,PIF1はDNAの二本鎖切断部位に集積し,G4DNAの位置で相同組換えを促進する46, 47).G4DNAは活性酸素種によって引き起こされる酸化的DNA損傷のセンサーとしても機能する.G4DNAの8-オキソグアニン修飾はプロモーターにおけるゲノム不安定性を引き起こす48).また,G4DNAの8-オキソグアニン修飾は,テロメアDNAの不安定性にも関与し49),テロメラーゼ活性を促進することが示されている50).
DNA複製におけるG4DNAの機能については,さまざまなモデルが提唱されている51).複製フォーク形成時,G4DNAは複製の障害物として機能する52).一方で,in silicoによるG4予測解析ではG4DNAが複製開始点に濃縮されていることから,G4DNAがDNA複製の開始を促進する可能性が示唆されている53).
転写におけるG4DNAの影響については複雑な細胞内メカニズムが関与しており,G4DNAの形成位置(鋳型鎖・非鋳型鎖・転写開始点の上流もしくは下流)によって異なる.鋳型鎖のG4DNAは,転写中のRNAポリメラーゼを直接阻害する可能性があり54, 55),非鋳型鎖のG4DNAは,新たに合成されたRNAとDNA/RNAハイブリッドG4を形成して転写終結を引き起こす56).転写開始点の上流でG4DNAが形成される場合,G4DNAは特定の転写因子をリクルートすることで転写を促進または抑制する57).転写とG4DNAの関連性に関しては,G4ChIP-seqによりG4DNA形成と転写活性に正の相関関係が報告されている30).プロモーターにおけるG4DNAは,細胞の分化過程におけるH3K4me3/H3K27me3バイバレント状態と関連しており,エピジェネティックな制御を介して細胞個性を形成する可能性がある58, 59).G4DNAはプロモーター領域のCpGアイランドのDNAメチル化を抑制し,遺伝子発現に寄与することも報告されている60).
前節で述べたとおり,G4ChIP-seqによって検出されたG4DNAのピーク数(約10,000か所)は,in silico予測によって検出された数(約300,000か所)やポリメラーゼ・ストップアッセイによって検出された数(約700,000か所)よりもかなり少ない.このことは,G4DNAの形成・解消が細胞内の何らかのメカニズムで調節されていることを示唆しているが,G4が特定の位置でいつ,どのように制御されているのかは未解明である.
2)G4RNAの生物学的機能
G4RNAはmRNAの5′および3′ UTRに豊富であり,遺伝子発現の調節に関与することが示唆される.G4RNAは,翻訳制御・選択的スプライシング・細胞内局在・RNA安定性など多様な生物学的機能に関与することが報告されているが,断片的な実験研究から推測されたものがほとんどであり,未解明な点が多い.
5′-UTRにおけるG4RNA(5′-G4RNA)は,無細胞および細胞におけるレポーターシステムを用いた実験で翻訳を抑制することが報告されている61–63).生体内の5′-G4RNAの翻訳制御メカニズムはまだ完全には解明されていないが,RNAヘリカーゼである翻訳開始因子eIF4Aの関与が報告されている.eIF4Aは,5′-UTRの核酸高次構造を解消し,43S開始前複合体をリクルートメントすることで翻訳を促進する.eIF4A阻害剤であるシルベストロールを添加したリボソームプロファイリングの結果,(CGG)4リピートからなる5′-G4RNAを持つmRNAの翻訳効率が低下した64).この結果は,G4RNAが43S開始前複合体のリクルートメントを抑制し,翻訳効率を低下させることを示唆している.翻訳阻害剤であるシクロヘキシミドを用いたHeLa細胞におけるリボソームプロファイリングでは,5′-G4RNAが上流オープンリーディングフレーム(upstream open reading frame:uORF)での80Sリボソーム形成を促進し,下流の主要なORFの翻訳阻害を引き起こすことが報告されている65).このメカニズムにはDEAHボックスRNAヘリカーゼであるDHX9およびDHX36が関与する.RNAヘリカーゼの種類によって5′-G4RNAの翻訳制御がどのように変化するのかを解明することは今後の課題となる.また,G4RNAが関与するuORFの翻訳メカニズムに関しては,後述するリピート伸長病の発症にも関与する.
ORFにおけるG4RNA(ORF-G4RNA)は,リボソームを一時停止しタンパク質分解やフレームシフトを誘導することが示唆されており,リボソームはORF-G4RNAの6~7ヌクレオチド手前で停止する66–69).また,レポーターアッセイにおいてORF-G4RNAは翻訳抑制の周期的な変動を3ヌクレオチドごとに引き起こすが,5′-G4RNAではみられない70).このORF-G4RNA特異的な現象が,実際に細胞内で観察されるかは興味深い.ORF-G4RNAとRNA結合タンパク質(RNA binding protein:RBP)との関連性についてもいくつか報告がある.APP mRNAのORF-G4RNAは,RBPであるFMRPと相互作用しmRNAの安定性に影響を与えることなく,翻訳を阻害する71).一方,MLL1 mRNAおよびMLL4 mRNAのORF-G4RNAは,RBPであるAVENおよびDHX36と相互作用し各mRNAのポリソーム会合を増加させ翻訳を促進する72).ORF-G4RNAとRBPの相互作用による翻訳伸長・リボソーム停滞・フレームシフトの制御メカニズムについても今後の研究成果が待たれる.
3′-UTRにおけるG4RNA(3′-G4RNA)は,5′-G4RNAと同様に翻訳を抑制的に調節することが知られている73, 74).一方で,3′-G4RNAは代替ポリアデニル化,miRNAターゲット相互作用,mRNA局在化などにも関与する.LRP5 mRNAとFXR1 mRNAの3′-G4RNAは,内部ポリアデニル化を促進し翻訳効率を上昇させる.特にFXR1 mRNAの場合,3′-UTRの短縮によりmiRNAとの相互作用が消失することで翻訳効率の上昇につながる75).ヒトの3′-G4RNA中の約44,000か所がmiRNA結合部位と重なることがバイオインフォマティクスで予測されている76).PSD-95 mRNAの3′-G4RNAはFMRPとmiR-125の両者が結合する.G4RNA形成時にはFMRPと相互作用し,一方でG4RNA解消時にはmiR-125とハイブリダイズする77).
神経細胞における3′-G4RNAは,軸索や樹状突起におけるmRNAの局所翻訳において重要な役割を担う.神経細胞のmRNA輸送は,シナプスで局所的に翻訳されシナプス可塑性と記憶の基盤となり,このプロセスの機能不全は神経機能を低下させる78–81).樹状突起mRNAは微小管に沿ってRBPやモータータンパク質とRNA顆粒を形成し輸送され,シナプスで局所的に翻訳が行われる.QGRS Mapperでの解析では,樹状突起mRNAの約30%が3′-G4RNAを持つ82).3′-G4RNAを持つ代表的な樹状突起mRNAとしてPSD-95 mRNAとCaMKIIα mRNAがあるが,どちらもG4RNA変異により樹状突起への輸送が阻害される82).興味深いことに,CaMKIIαの3′-UTRが欠損した遺伝子変異マウスでは,CaMKIIα mRNAの樹状突起への輸送が低下し記憶学習の低下を示す83)が,3′-G4RNAがどの程度関与するのかは未解明である.
本稿では割愛するが,ノンコーディングRNA(lncRNA, miRNA, piRNA, tRNA, rRNA)やRNAウイルスにおけるG4RNAの存在と役割も報告され始めている84).しかしながら,これまでの細胞におけるG4RNA研究のほとんどは,内在性G4RNAを必ずしも再現しないレポーターアッセイを用いた解析である.今後,細胞内におけるG4RNAの生物学的機能を明らかするためには,内在性RNA構造を反映した包括的なトランスクリプトーム解析法が必要である.
これまで,グアニンリッチ配列の伸長によるリピート伸長病においてG4の異常形成が神経疾患の発症に関与することが示唆されている.中でも,C9ORF72遺伝子変異を起因とする筋萎縮性側索硬化症・前頭側頭葉変性症(C9ALS/FTD)の研究が最も盛んに行われている.C9ALS/FTDでは,C9ORF72遺伝子非翻訳領域内にヘキサヌクレオチドGGGGCC(G4C2)リピート伸長が起こり,DNAおよびRNAでG4とヘアピン構造が混在した状態が形成される85, 86).この核酸構造異常に起因した病態メカニズムがこれまでに四つ提唱されている87).①RNA毒性:G4C2リピートのRNA転写産物が核内にRNA凝集体を形成し,その凝集体に多くのRBP群を巻き込み,それらを機能不全にすることで神経変性を引き起こす.②Repeat-Associated Non-AUG(RAN)翻訳:リピート伸長RNAにおいて,すべてのリーディングフレームでジペプチドリピートタンパク質が異常翻訳され,神経変性を引き起こす.③C9ORF72タンパク質ハプロ不全:G4DNAがRNAポリメラーゼを失速させ,C9ORF72タンパク質産生を抑制する.C9ORF72タンパク質の減少は,グルタミン酸受容体の凝集・機能不全を誘導し神経変性を引き起こす.④内因性DNA損傷:G4DNAが二本鎖切断の形成を促進し,DNA修復経路の機能不全を引き起こす.その他の疾患例として,脆弱X随伴振戦/失調症候群(fragile X-associated tremor/ataxia syndrome:FXTAS)があげられる88).FXTASでは,FMR1遺伝子の5′非翻訳領域におけるCGGリピート伸長がみられる.FXTASでは,C9ALS/FTDと同様にRNA毒性が観察され,RNA代謝に異常を引き起こす.またRNA毒性に加えて,RAN翻訳も観察される.伸長したCGGリピートRNAは,RAN翻訳を介して有毒なポリグリシン含有タンパク質「FMRpolyG」を産生する89).
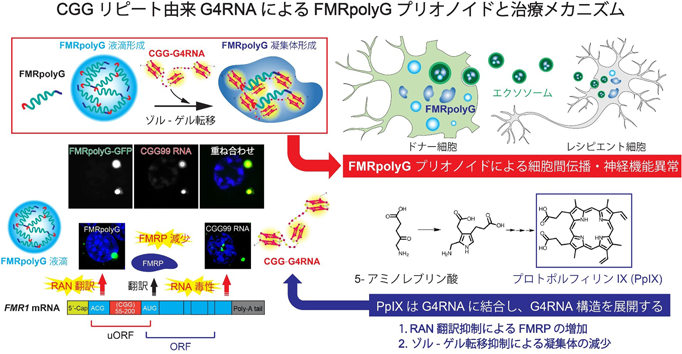
筆者らは,FMRpolyGに含まれるCGGリピート由来のポリグリシン領域にプリオン様の性質があることに着目した.プリオン様領域を持つタンパク質は液–液相分離(liquid-liquid phase separation:LLPS)により液滴を形成することが知られている.予想どおり,FMRpolyG精製タンパク質は液滴を形成した.また,FMRpolyGの液滴をCGG99リピート(FXTAS発症リピート数)RNAと混合して観察したところ,FMRpolyGはCGG99リピートRNAと複合体を形成しゲル状の凝集体となった.RNAの物性解析を行ったところ,健常人のリピート数であるCGG17リピートRNAがヘアピン型であるのに対し,CGG99リピートRNAはG4構造を形成すること,また,CGG99リピートRNAはFMRpolyGのポリグリシン領域と直接結合することを見いだした.つまり,FMRpolyGはポリグリシン領域を介して,G4RNAと結合することでゾル–ゲル相転移することが明らかになった(図4)90).
次に,FMRpolyGに結合するタンパク質を網羅的に解析した.その結果,FMRpolyGは,神経変性疾患の発症に関与するRBP群(HNRNPA2/B1, FUS, SFPQなど)と結合することがわかった.また,FMRpolyGは細胞外小胞であるエクソソームに含まれるタンパク質群とも複合体を形成することを見いだした.FXTASモデルマウスの脳由来エクソソームにはFMRpolyGが高発現しており,野生型マウス神経細胞(レシピエント細胞)にFXTASモデルマウス神経細胞(ドナー細胞)由来エクソソームを処置することによって,野生型マウス神経細胞にもFMRpolyGの発現が確認され,神経機能異常がみられた.つまり,FMRpolyGはエクソソームを介して細胞間伝播し,神経機能異常を引き起こす「プリオノイドタンパク質」であることを発見した.さらに,生体内ポルフィリンであるプロトポルフィリンIX(proto-porphyrin IX:PPIX)が,G4に結合することでプリオノイドタンパク質の凝集を抑制することを発見した.PPIXをFMRpolyGとG4RNAの複合体に処置することで,LLPSを介した凝集体形成が劇的に抑制された.体内でPPIXを産生する5-アミノレブリン酸をFXTASモデルマウスに経口投与することでRAN翻訳を抑制し,神経伝達機能・認知機能・運動機能の低下を有意に改善することができた(図4)90).
近年,アルツハイマー病やパーキンソン病を代表するタウオパチーやシヌクレイノパチーの発症にプリオノイド機構が関与することが注目されているが,その細胞内メカニズムは未解明である.FXTASの結果から,プリオノイドの引き金がG4RNAであると考えられた.本稿では割愛するが,筆者らは,タウやαシヌクレインの凝集がG4RNAにより引き起こされることを明らかにした91, 92).さらに近年,FXTASと同様の神経症状を呈するグアニンリッチ・リピートに由来する神経変性疾患が近年のロングリード・シーケンス技術により多数発見されている.筆者らが発見したプリオノイド機構「G4プリオノイド」を基盤とした神経変性疾患全般(遺伝性および孤発性)に対する病態解明と治療薬開発が期待される93).