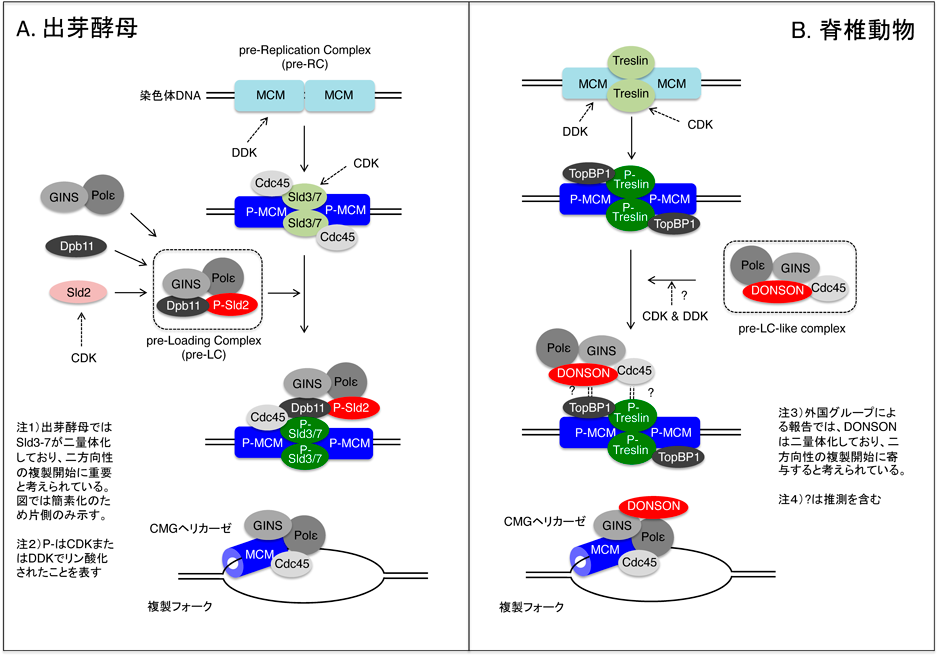
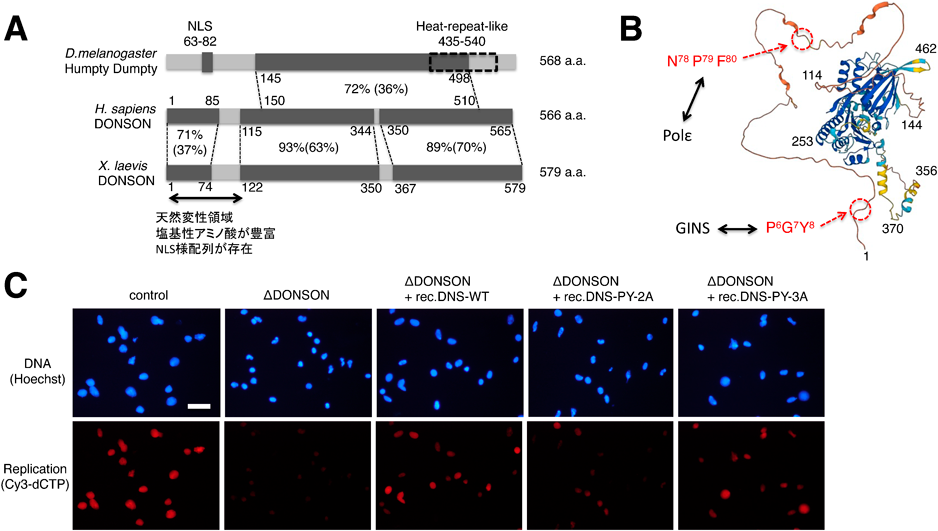
高等真核生物におけるDNA複製開始機構Mechanism of DNA replication initiation in higher eukaryote
東京薬科大学生命科学部Tokyo University of Pharmacy and Life Sciences ◇ 〒192–0392 東京都八王子市堀之内1432–1 ◇ 1432–1 Horinouchi, Hachioji-shi, Tokyo 192–0392, Japan
発行日:2024年12月25日Published: December 25, 2024