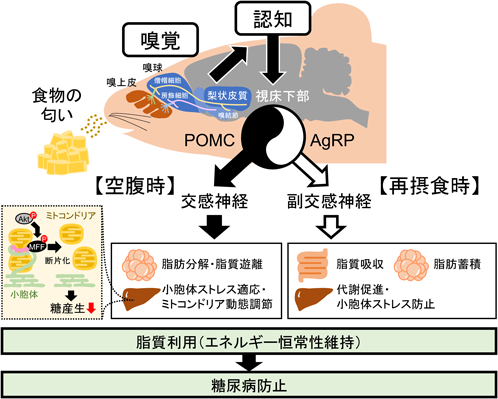
嗅覚と代謝:認知によってつながる関係Olfaction and metabolism: How are they connected?
富山大学学術研究部(薬学・和漢系)臨床薬品作用学研究室Department of Integrative Pharmacology, University of Toyama ◇ 〒930–0194 富山市杉谷2630 ◇ 2630 Sugitani, Toyama 930–0194, Japan
発行日:2025年2月25日Published: February 25, 2025