皮膚の感覚神経の細胞体は,脊髄の横に並ぶ後根神経節や,脳幹の側にある三叉神経節に存在する1, 2).これらの感覚神経細胞は,二股に分かれた軸索を持ち,片方は皮膚に,もう片方は中枢神経系に投射している.後根神経節の感覚神経細胞は,後頭部および首から下の皮膚からの信号を脊髄に伝え,三叉神経節の感覚神経は,前頭部および顔の皮膚からの信号を脳幹へと伝える.近年,個々の細胞のRNA発現を網羅的に解析するシングルセルトランスクリプトーム解析が,さまざまな組織細胞について行われるようになり,後根神経節や三叉神経節の感覚神経細胞についても,マウスだけでなくヒトでもすでに複数の報告がある2–4).遺伝子発現の特徴から,個々の感覚神経細胞が,有髄神経または無髄神経のどちらを形成するか,そしてどのような機能を持つか,ある程度推測することができる.マウスの後根神経節の感覚神経細胞は,遺伝子発現パターンによって,大まかに11~12種類のサブセットに分かれると報告されている2–4).また,ヒトの後根神経節の感覚神経細胞は,大まかに11~16種類の遺伝子発現パターンに分かれると報告されている2–4).もちろん,これらすべてが皮膚に投射しているわけではなく,一部は,体の部位の位置や運動感覚を伝える固有受容器として,筋肉,腱,関節などに投射しており,また腸などの体内組織にも投射している.皮膚に投射する神経のうち,痛みや痒みを誘導する神経線維の多くは無髄神経であるC線維であり,皮膚における神経線維の末端は特別な構造がみられない自由神経終末となっている1, 2).これらの神経は侵害受容器と呼ばれ,古典的にはひとくくりにされており,その多様性は明らかではなかった.一方,触覚や圧覚を伝える神経線維の多くは有髄神経であるAβ線維であり,毛包の周りを取り囲んでいたり,メルケル盤やマイスナー小体などの構造体の一部となっていたりと,多様な神経終末構造を持つ1, 2).
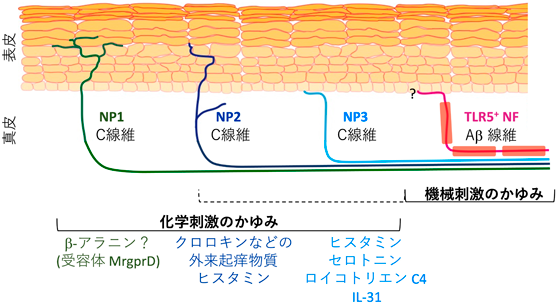
マウスの後根神経節では,11~12種類の感覚神経細胞サブセットのうち,3種類のサブセットに,痒みを惹起する物質の受容体の遺伝子が発現していることが示された2–4).異なる呼称を使っている報告もあるが,非ペプチド作動性(non-peptidergic:NP)のNP1, NP2, NP3という呼称が,この3種類のサブセットに対して現時点では最もよく使われている.ただ,この呼称には問題もある.侵害受容器のうち,カルシトニン遺伝子関連ペプチド(calcitonin gene related peptide:CGRP)やサブスタンスPといった神経ペプチドを放出する神経をペプチド作動性(peptidergic:PEP),それ以外の侵害受容器を非ペプチド作動性としているが,実際にはNP2もCGRPを発現しており,またその他にもNP1, NP2, NP3に発現する神経ペプチドやペプチドホルモンが存在する.そしてこの問題は,後に述べるとおり,ヒトの感覚神経サブセットの解析結果でも顕著となっている.そのため,いずれは別の呼称に改められる可能性もあるが,ここではこの呼称を使うこととする.NP1, NP2, NP3神経の特徴については,筆者が以前,別の稿でまとめているので,それを引用し,その後のアップデートを加えて述べる2, 5).
NP1神経サブセットには,ヒスタミンのような古典的な内因性起痒物質の受容体は発現していないが,Mas-related G-protein-coupled receptor(Mrgpr)ファミリーの一つであるMrgprDが特異的に強く発現している.MrgprDは起痒物質であるβ-アラニンの受容体であり,MRGPRDを欠損したマウスは,β-アラニン投与による痒みが消失すると報告されている2, 5).したがって,β-アラニン刺激による痒みの伝達は,NP1によって担われていると考えられる.しかしながら,MrgprDおよびこれを発現する神経は,痛み,温度,機械刺激の感覚にも関わるという報告もある2, 5).よってNP1は痒み,痛み,温度などの感覚を伝達する多モード侵害受容器であると考えられている.NP1のほとんどの神経終末が,皮膚の表皮層の最も表面近くまで伸びると考えられている(図1)2, 5, 6).
NP2神経サブセットには,ヒスタミンの受容体の発現が検出されただけでなく,MrgprファミリーのMrgprA3が選択的に発現している.抗マラリア剤などとして用いられていたクロロキンは,その副作用の一つとして痒みを誘導することが知られており,これはMrgprA3に直接作用する結果だと報告されている2, 5).MrgprA3を発現する神経だけを欠失させると,さまざまな起痒物質による痒みが減弱するだけでなく,乾燥皮膚モデルや接触性皮膚炎モデルでの痒みも減弱すると報告されている2, 5, 7).これらの結果から,NP2は痒みの伝達に特化した神経であると提唱されている8).NP2神経の終末も,NP1と同様に,表皮層の最も表面近くまで伸びると報告されているが,NP1の場合と異なり,どのくらいの割合のNP2神経線維が,表皮層に入っているか明らかではない(図1)2, 5, 7).
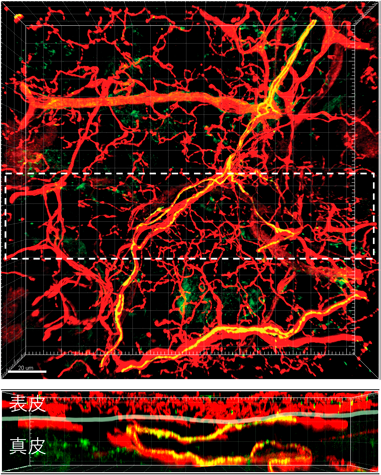
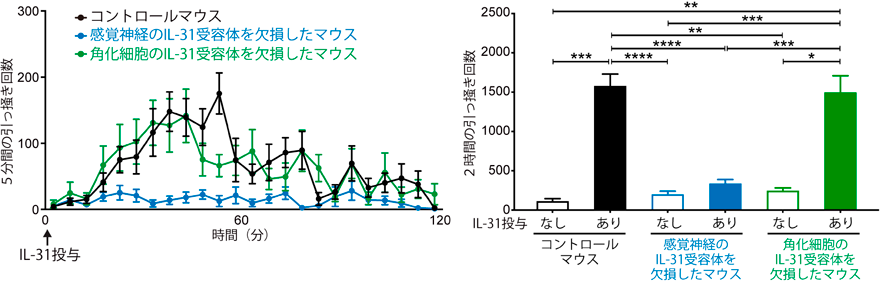
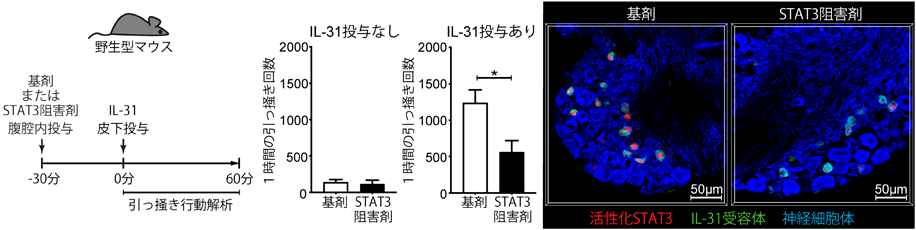
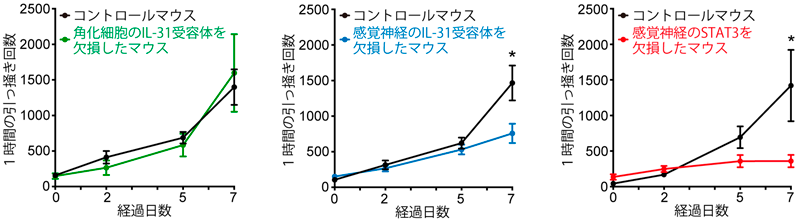
NP3神経サブセットには,ヒスタミンやセロトニンといった,主に肥満細胞が放出することが知られる起痒物質の受容体が発現するだけでなく,痒みを誘導するサイトカインIL-31の受容体が選択的に強く発現する2, 5).筆者らの最近の研究によって,NP3神経線維の終末は,マウスの健常皮膚ではほとんど表皮に入っておらず,真皮内または真皮と表皮の境界に存在していることが明らかとなった(図1,図2)2, 9).ただし,皮膚炎の状態では,NP3神経終末も,表皮に入っていく可能性もある(未発表データ).IL-31受容体を感覚神経特異的に欠損させたマウスでは,IL-31投与によって惹起される痒みが消失することもわかり,IL-31は直接神経に作用して痒みを惹起していることが証明された(図3)2, 9).それに対して,角化細胞特異的にIL-31受容体を欠損させても,IL-31が惹起する痒みは減弱しなかった(図3)2, 9).
NP1, NP2, NP3神経は,上記に加えて,IL-31と同様に2型サイトカインであるIL-4やIL-13の受容体の遺伝子を,弱いながらも発現している2, 5).IL-4やIL-13は皮下に投与しても,それ自体だけでは強い痒みを引き起こしにくいが,IL-31やほかの起痒物質を低容量で投与したときの痒みを,著しく増強することが報告された2, 5, 10).また,IL-4やIL-13の受容体を感覚神経特異的に欠損させたマウスでは,他の起痒物質の痒みの増強効果や,皮膚炎モデルの痒みが抑制されることも示されている2, 5, 10).
3. IL-31の痒みにおける感覚神経STAT3の役割
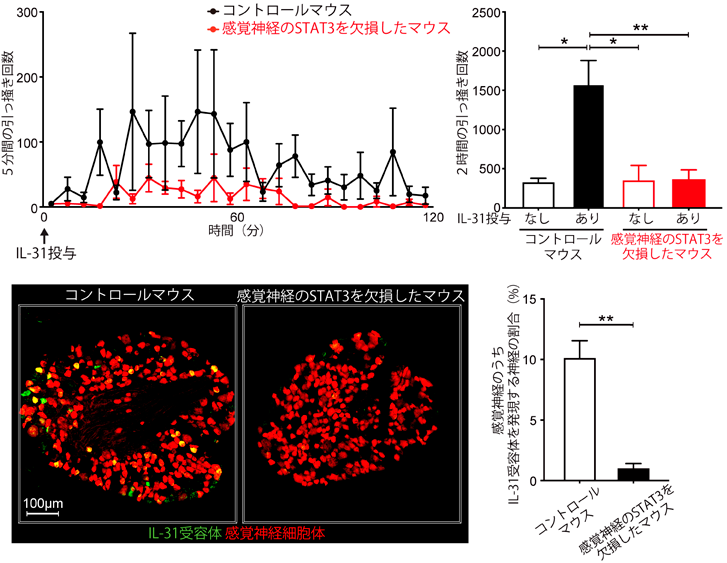
IL-4, IL-13, IL-31といったサイトカインシグナルが,どのように感覚神経を活性化するのかについては,まだまだ謎が多い.これらの受容体直下ではヤヌスキナーゼ(JAK)が活性化され,これらのサイトカインによる痒みにおいても,感覚神経のJAKが重要であることは報告されていた10).JAKの下流ではSTATファミリーの活性化が起こり,細胞核の中へと移行して転写因子として働く.免疫細胞をはじめ多くの細胞のサイトカイン応答において,STATファミリーによる遺伝子発現誘導が,重要な役割を果たすことが知られている.しかしながら,痒み誘導における感覚神経のSTATファミリーの役割は,限定的なのではないかと考えられていた.後根神経節から単離した感覚神経を,上記のサイトカインで刺激すると,活性化のイベントの一つである細胞内カルシウム上昇が起こるが,これはサイトカイン刺激後の数分以内に起こりうることが報告され,これにSTATファミリーによる遺伝子発現誘導が関与するのは,時間的に考えにくいと思われた10).しかしながら,図3にも示すように,IL-31投与後,マウスの引っ掻き行動は15分以上経過してから起こるため,筆者らは,痒み誘導にSTATファミリーによる遺伝子発現誘導が関与しうるかもしれないと考えた.
シングルセルトランスクリプトーム解析の結果から,NP1, NP2, NP3を含めたほとんどの種類の感覚神経において,最も顕著に発現しているSTATファミリー分子は,STAT3であることを見いだした9).そこで,STAT3を感覚神経においてのみ欠損させたマウスを新たに作製した.このマウスに,IL-31を皮下投与すると,IL-31が惹起する引っ掻き行動がまったく観察されなかった(図4上段)9).この原因を調べるため,まず,STAT3欠損により感覚神経の痒み関連分子の発現が変化していないか調べたところ,IL-31受容体の発現が減弱しているという予期せぬ結果を得た(図4下段).つまり,感覚神経がIL-31受容体を発現するメカニズムに,STAT3が深く関わっていることが明らかとなった.
上記のように,遺伝子改変によって,感覚神経のSTAT3をマウスの発生途中から欠損させてしまうと,IL-31受容体の発現を低下させることがわかったため,感覚神経のSTAT3が,IL-31受容体下流の痒み誘導シグナル伝達に関わっているかどうかは解析できなかった.そこで,STAT3活性化に対する小分子阻害剤を,野生型マウスに投与することで,IL-31が誘導する痒みに影響が出るかを調べた.その結果,阻害剤投与により,IL-31による感覚神経のSTAT3活性化が減弱し,IL-31が惹起する引っ掻き行動が,消失はしないものの有意に減弱した(図5)9).一方,阻害剤投与によるIL-31受容体の発現低下は確認されなかった(図5)9).以上のことから,STAT3はIL-31受容体下流の痒み誘導シグナル伝達に関わっていることが示唆された.
さらに,皮膚炎の痒みにおけるIL-31受容体や感覚神経STAT3の役割について,解析を行った.アトピー性皮膚炎に似た2型炎症を起こす皮膚炎モデルを,角化細胞においてのみIL-31受容体を欠損させたマウスに適用したところ,皮膚炎に伴う引っ掻き行動は,コントロールマウスと同程度観察され,皮膚炎モデルにおいても,角化細胞のIL-31受容体の重要性は認められなかった(図6)9).しかしながら,感覚神経においてのみIL-31受容体を欠損させたマウスに適用したところ,皮膚炎に伴う引っ掻き行動が減弱した.この減弱は有意ではあるものの部分的なものであり,引っ掻き行動はまだはっきりと残っていた(図6)9).よって,この皮膚炎モデルにおける痒み誘導においても,感覚神経のIL-31受容体の関与が認められたものの,感覚神経のIL-31受容体が関与しない痒みも存在することが示唆された.一方,感覚神経においてのみSTAT3を欠損させたマウスに,この皮膚炎モデルを適用したところ,引っ掻き行動が強く抑制されていた(図6)9).よって,感覚神経のSTAT3は,IL-31が惹起する痒みだけでなく,皮膚炎においてほかのメディエーターが惹起する痒みにも重要な役割を果たしていることが示唆された.今後は,感覚神経においてSTAT3依存的に発現誘導される遺伝子のなかで,痒みに重要な役割を果たすものを探索していく必要がある.
4. 機械刺激が誘導する痒みを伝達するマウス感覚神経
くすぐるような機械刺激を受けると,くすぐったいのとともに,痒みが誘導される場合がある.この機械刺激による痒みは,従来は触覚や圧覚を伝えるとされた有髄Aβ線維神経である低閾値機械受容器(low threshold mechanoreceptor:LTMR)のうち,Toll-like receptor 5(TLR5)の遺伝子を発現するサブセットによって伝達されると報告された2, 5, 11).ただ,Tlr5発現が,機械刺激の痒みを伝達する神経サブセットに,どのくらい特異的なのかはまだわかっていない.またこの神経サブセットの皮膚内分布や終末構造も不明である.同じ報告では,NP1, NP2, NP3を含むすべての無髄感覚神経を除去できる遺伝子改変マウスが作製され,このマウスでは化学刺激による痒みは消失するものの,機械刺激による痒みは減弱しないというデータも示された11).筆者らも,類似の遺伝子改変マウスを作製して実験したところ,上記と一致する結果を得た(未発表データ).しかしながら,その後,NP3神経は,Piezo1という機械刺激感受性イオンチャネルを発現し,これによって機械刺激による痒みを誘導・伝達するという報告も出た12).そして,ヒスタミンなどがNP3に作用すると,NP3の伝達する機械刺激の痒みを生じやすくすると唱えられた.さらに,NP2神経も,Piezo2という機械刺激感受性イオンチャネルを発現し,ヒスタミンなどが増強する機械刺激の痒みの伝達に関わっているとも報告された13).よって,機械刺激の痒みを伝達する神経が,有髄Aβ線維なのか無髄C線維なのか,それとも両方なのかはまだ決着がついていない.ただ,上述の筆者らによる追試の結果から,有髄神経の貢献の方が大きいのではないかと,現時点では筆者は考えている(図1).
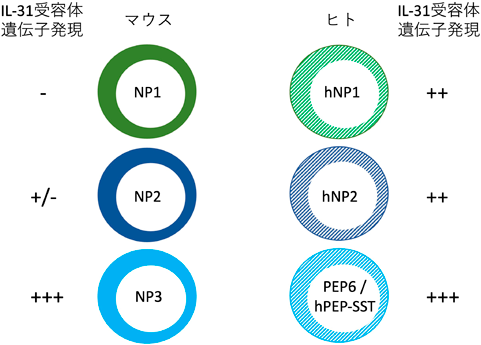
ヒト後根神経節のシングルセルトランスクリプトーム解析も,すでに複数の報告があり,マウスのNP1, NP2, NP3神経の遺伝子発現パターンと類似点を持つ感覚神経サブセットが見いだされている.2021年に出た報告では,ヒスタミンやIL-31などの痒み誘導物質の受容体の遺伝子を発現するサブセットは二つしかないとされた2, 3, 14).そして,その二つともがIL-31受容体遺伝子を発現しており,マウスにおけるNP3サブセットの特徴を持っているとされた.それと同時にこの二つのサブセットは,マウスにおけるNP1やNP2サブセットの遺伝子発現の特徴も,互いに異なる形で有しているようであった2, 3, 14).一方,その後の複数の報告では,痒み誘導物質の受容体の遺伝子を発現する感覚神経サブセットは,ヒトでも三つ存在し,NP1, NP2, NP3との対応づけも可能な程度に,遺伝子発現パターンの類似性があると報告された2, 3, 13).もちろんマウスと異なる点も多くあり,やはりIL-31受容体の遺伝子発現は,NP3に対応するサブセットに限局しておらず,三つのサブセットすべてに発現しているとされた(図7).またIL-31受容体遺伝子だけでなく,マウス感覚神経においてサブセット特異的または選択的な発現を示す遺伝子が,ヒト感覚神経サブセットでは,その特異性が弱く,多くのサブセットにまたがって発現している傾向が示された2–4, 15).なかでも,ペプチド作動性(PEP)を特徴づけるCGRPの遺伝子発現は,ほとんどすべてのサブセットに発現しているという報告もある2, 4).さらに,NP3に対応するとされたヒト感覚神経は,ペプチド作動性(PEP)のサブセットの一種とする報告もある2–4, 15).上述のように,マウスにおいてもNP1, NP2, NP3を非ペプチド作動性(NP)とするのは,実際の性質と齟齬がある可能性があり,混乱を招かない新たな呼称が必要と思われる.ヒトにおいて,機械刺激が誘導する痒みを伝達する感覚神経サブセットについては,残念ながらまだ知見がほとんどない.