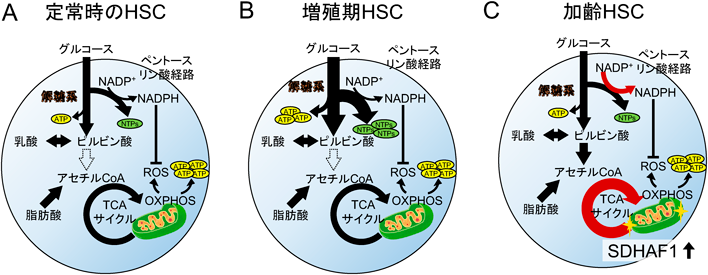
単一細胞代謝解析技術が見いだした造血幹細胞の代謝柔軟性Metabolic flexibility of hematopoietic stem cells
1 東北大学大学院医学系研究科幹細胞医学分野Department Cell Fate Biology and Stem Cell Medicine, Tohoku University Graduate School of Medicine ◇ 〒980–8575 宮城県仙台市青葉区星陵町2–1 ◇ 2–1 Seiryo-machi, Aoba-ku, Sendai 980–8575, Japan
2 国立健康危機管理研究機構国立国際医療研究所造血システム研究部Department of Stem Cell Biology, National Institute of Global Health and Medicine, Japan Institute for Health Security ◇ 〒162–8655 東京都新宿区戸山1–21–1 ◇ 1–21–1 Toyama, Shinjuku-ku, Tokyo 162–8655, Japan
発行日:2025年6月25日Published: June 25, 2025