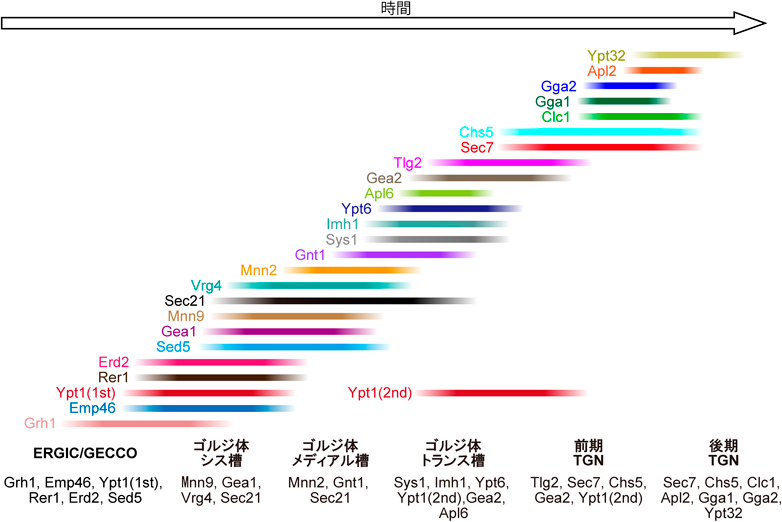
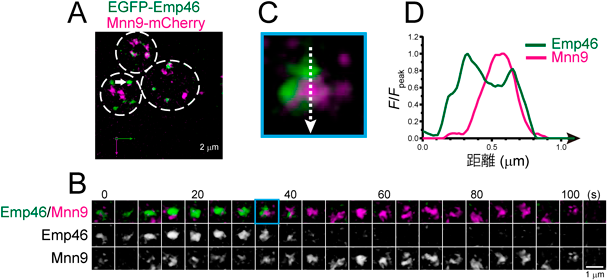
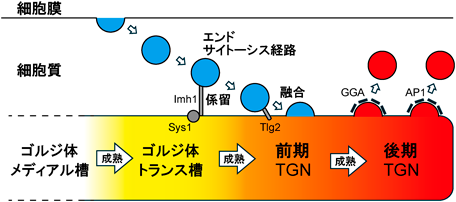
超解像ライブイメージングが解き明かすゴルジ体槽成熟の時空間ダイナミクスSpatiotemporal dynamics of Golgi cisternal maturation revealed by super-resolution live imaging
理化学研究所光量子工学研究センター生細胞超解像イメージング研究チームLive Cell Super-resolution Imaging Research Team, RIKEN Center for Advanced Photonics ◇ 〒351–0198 埼玉県和光市広沢2–1 ◇ 2–1 Hirosawa, Wako, Saitama 351–0198, Japan
発行日:2025年6月25日Published: June 25, 2025