ARLファミリー低分子量Gタンパク質の多様な機能Functional diversity of ARL family small GTPases
明治薬科大学薬学部Meiji Pharmaceutical University ◇ 〒204–8588 東京都清瀬市野塩2–522–1 ◇ 2–522–1 Noshio, Kiyose, Tokyo 204–8588
発行日:2025年8月25日Published: August 25, 2025
150種類以上知られている低分子量Gタンパク質のうち,ARF/ARLファミリーに属する分子はヒトでは29種類存在する.このファミリー中で,SAR1やARFについてはこれまで精力的に解析が進められ,小胞体やゴルジ体において物質輸送を制御することなどが明らかにされてきた.一方,ARF/ARLファミリーの半数以上を占めるARL(ARF-like)については,それらの多くがARFとの相同性に基づきデータベース上で同定されてきた経緯もあり,機能解析が遅れている.しかし近年,さまざまな研究によってARLに関する解析が進み,それらが一般的なARFと比べて,局在する部位や制御する細胞応答などの点で非常に多様性に富むことがわかってきた.本稿では各種のARLに関して,最近までにわかってきた知見や今後の展望などについて紹介したい.
© 2025 公益社団法人日本生化学会© 2025 The Japanese Biochemical Society
低分子量Gタンパク質は一次構造上の特徴から,RAS, RHO/RAC, RAB, ARF/ARL, RANファミリーに分類される.このうちARF/ARLファミリーに分類される分子は,ヒトでは29種類存在し,SAR1(2種類),ARF(5種類),ARL(20種類),ARFRP1, TRIM23,から構成される1).ARF/ARLファミリーの中で半数以上を占めるARLは,主にARFとの相同性からデータベース上で同定されてきたものが多く,真核生物で広く保存されている分子もあれば,進化的に比較的新しく出現したと思われる分子も存在する.現在までにわかってきたARLの特徴の一つとしては,その多様な細胞内局在があげられる.ARFは主にゴルジ体に局在して機能しているが,ARLはトランスゴルジ網(trans-Golgi network:TGN)に加え,一次繊毛,ミトコンドリア,リソソーム,核,中心体,など非常に多様な場所に局在しており(図1),結果として多岐にわたる細胞応答の制御に関与している.
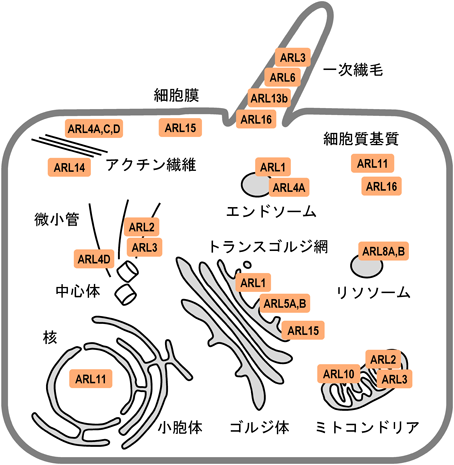
各ARLが主に局在すると考えられる細胞内の部位を示した.ARL3のように,複数部位に局在すると思われる分子も存在する.
ARL分子群の中には,ARFにはみられない特徴的な一次構造を有する分子も存在する.一般的なARFでは,N末端から2番目のグリシン残基のミリストイル化修飾が膜局在化に必要なのに対し,ある種のARLではN末端近傍のシステイン残基のパルミトイル化修飾が膜局在化に必要であり,N末端領域に膜貫通ドメインを有するARLも存在する.さらに,低分子量Gタンパク質としてのドメインに加えて,別の機能ドメインを有するARLも存在する.本稿では,各種のARLに関して,構造上の特徴も含め,最近までにわかってきた知見を以下に御紹介したい.
酵母から哺乳動物までよく保存されており,主にTGNに局在し,多様な細胞応答に関与している2).TGNでは,Golgin-245やGolgin-97といった係留タンパク質をリクルートすることで,エンドソームからTGNへの逆行性輸送に関与すると考えられている3, 4).ARL1のTGNへの局在化には,ARF/ARLファミリーの一つであるARFRP1が関与すると考えられている5).またARL1はソーティングエンドソームにも局在し,ARFの活性化因子であるBIG1をリクルートして,ARF1やARF3の活性化にも関与することが報告されている6).
細胞内のさまざまな部位(細胞質基質,ミトコンドリア膜間部,中心体,核など)に存在することが報告されており,αβ-チューブリンの生合成7),ミトコンドリアの融合8, 9),中心体からの微小管伸張10, 11)などに関与することが報告されている.
ARL2と相同性が高く,ARL2と同様の機能を有すると考えられているが,脂質修飾タンパク質の一次繊毛への局在化にも関与することが知られている(図2)12, 13).細胞小器官(オルガネラ)の一つである一次繊毛は,ヒトでは多くの組織の細胞に存在し,外界環境のセンサーとして機能している.一次繊毛の膜に局在するタンパク質の中で,ミリストイル化修飾やプレニル化修飾がその膜局在に寄与するタンパク質は,それらの脂質部分を認識するシャペロン様タンパク質であるUNC119やPDE6Dと,それぞれ結合した状態で一次繊毛へと輸送される.その後,一次繊毛内において,GTP結合型のARL3がUNC119やPDE6Dと結合することで,脂質修飾タンパク質が遊離して一次繊毛の膜に局在化するようになると考えられている.興味深いことに,ARL分子群の一つであるARL13B(後述)とBARTと呼ばれる因子が協調的にARL3のGEF(guanine nucleotide exchange factor)として働くことで,GTP結合型のARL3が生成すると考えられている14–16).一方,GTP結合型のARL3は,ARL3のGAP(GTPase-activating protein)であるRP2の作用により,GDP結合型になると考えられている17–19).
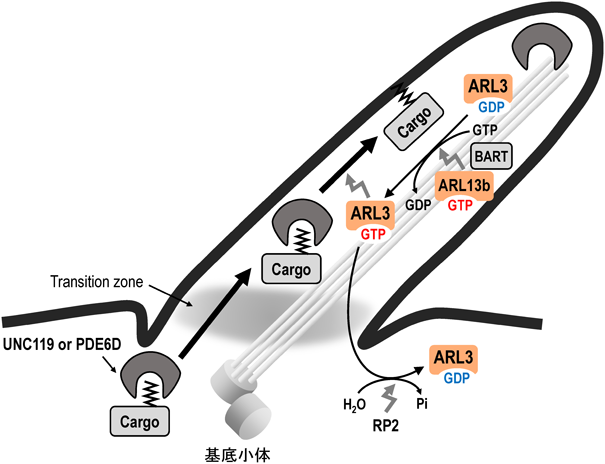
ミリストイル化修飾あるいはプレニル化修飾された一次繊毛局在性タンパク質は,シャペロン様タンパク質UNC119あるいはPDE6Dと結合して繊毛内に輸送される.一次繊毛内では,ARL13BとBARTが協調的にARL3のGEFとして機能することで,GTP結合型のARL3が生成する.GTP結合型のARL3は,UNC119やPDE6Dに作用し,それらと複合体を形成していた脂質修飾タンパク質は解離し,一次繊毛の膜に局在するようになる.GTP結合型のARL3は,繊毛外において,ARL3-GAPであるRP2の作用により,GDP結合型に変換される.
細胞内のさまざまな部位(細胞膜,エンドソーム,ゴルジ体,核)に存在することが報告されている.エフェクタータンパク質として,ELMO(DOCK180と協調して低分子量Gタンパク質RACの活性化を制御)やRobo1が報告されており,アクチン骨格のリモデリングや細胞移動に関与することが示唆されている20, 21).ゴルジ体においては,係留タンパク質であるGCC185と相互作用することで,エンドソーム-ゴルジ体間の輸送に関与することが示唆されている22).また,エンドソームにおいてタンパク質のソーティングに重要なESCRT複合体のサブユニットであるVPS36と相互作用し,EGF受容体の分解に関与することが示唆されている23).
相互作用因子として,ARF6のGEFであるCytohesin-2/ARNO, RAC1およびCDC42の活性化に関与するIQGAP1,アクチン結合タンパク質Filamin-Aが知られており,アクチン骨格のリモデリングや細胞移動に関与すると考えられている24–26).ARL4Cの遺伝子発現は,Wnt/β-CateninやEGF/RASシグナル伝達経路によって調節されると考えられており,いくつかのがんでその発現上昇がみられることから,がんの浸潤などとの関連性が指摘されている27–31).
Cytohesin-2/ARNOと相互作用することで,アクチン骨格のリモデリングに関与することが示唆されている24, 32, 33).またARL4DのGTP結合型は,EB1(伸張する微小管の先端に結合するタンパク質)の中心体への集積を促進し,EB1とダイナクチンサブユニットp150の相互作用を促進することで,微小管の核形成と成長を制御することが示唆されている34).
ARL5AおよびARL5BはTGNに局在し,GARP(Golgi-associated retrograde protein)をTGNにリクルートすることで,エンドソームからゴルジ体への逆行性輸送を制御すると考えられている35, 36).またARL5Bに関しては,ACBD3(acyl-CoA-binding domain-containing 3)のTGNへの局在化にも関与することが報告されている37).ARL5BのTGNへの局在化には,ARFRP1が関与することが報告されており5),シャペロンタンパク質HSC70がGDP結合型のARL5Bと相互作用し,ARL5BのTGNへの局在化に関与することも示唆されている38).現時点でARL5Cに関する知見は報告されていない.
一次繊毛の形成・機能に関与することが知られており,繊毛性疾患バルデー・ビードル症候群(Bardet–Biedl syndrome)の原因遺伝子の一つである(遺伝子名:BBS3).一次繊毛が外界環境を感知するセンサーとして機能するためには,ある種の受容体やチャネルが一次繊毛に局在することが必要であるが,それらの一次繊毛への輸送にはBBSomeと呼ばれるタンパク質複合体が関与する.また一次繊毛内においては,BBSomeに加えて,IFT(intraflagellar transport)と呼ばれるタンパク質複合体が介在する.IFTは,IFT-A(6個のサブユニット)とIFT-B(16個のサブユニット)から構成されており,IFT-Aはダイニンモーターと相互作用して繊毛内での逆行性輸送を,IFT-Bはキネシンモーターと相互作用して順行性輸送を担っている39, 40).
ARL6のGTP結合型は,BBSomeと結合することが示されており41),その結合にはIFT22(RABL5)が関与することが報告されている(IFT22はIFT-Bのコンポーネントであり,低分子量Gタンパク質の一つ)42).細胞質においてARL6とBBSomeからなる複合体は,IFT22(RABL5)と結合することで基底小体に輸送され,その後IFT22がその複合体から解離し,ARL6-BBSome複合体がIFT複合体によって繊毛内で輸送されるモデルが提唱されている(図3).バルデー・ビードル症候群で同定されているARL6の変異は,ARL6タンパク質の安定性やGTPに対する結合親和性の低下を引き起こすことが知られている43).
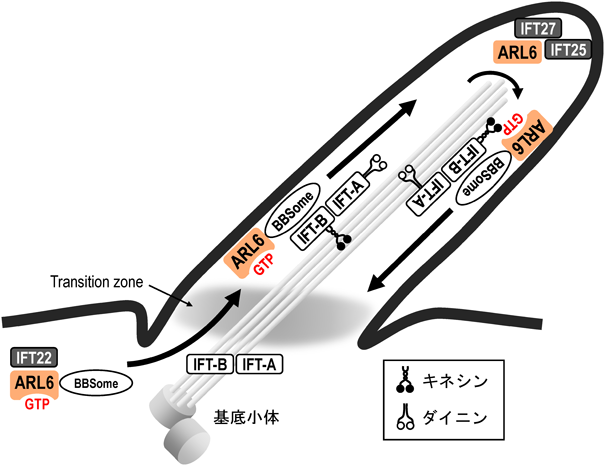
BBSomeは一次繊毛への膜タンパク質輸送に介在するタンパク質複合体であり,GTP結合型ARL6はBBSomeと結合し,IFT22の作用により基底小体に運ばれる.その後,ARL6-BBSome複合体はIFT22から解離し,IFT複合体によって繊毛内へ輸送される.繊毛内においてARL6-BBSome複合体は,IFT-Bに結合したキネシンモーターの働きによって順行性に輸送されるが,頂端部においてIFT複合体の解離と再会合が生じ,IFT-Aに結合したダイニンモーターの働きによって逆行性に輸送されるようになる.頂端部においては,IFT27(RABL4)とIFT25から構成されるヘテロ二量体が,グアニンヌクレオチドを結合していないARL6と結合し,上記の輸送の方向転換に寄与することが示唆されている.
繊毛内に輸送された膜タンパク質は,順行性輸送により一次繊毛内を輸送されるが,一次繊毛の頂端において,IFT複合体の解離と再会合が生じ,順行性輸送から逆行性性輸送への転換が起こり,繊毛外へ輸送されると考えられている.一次繊毛の頂端においては,IFT27(RABL4)とIFT25から構成されるヘテロ二量体が,グアニンヌクレオチドを結合していないARL6と相互作用することが示唆されており,両者の相互作用が標的タンパク質の繊毛外への輸送に関与することが示唆されている44, 45).
主にリソソームに局在し,微小管依存的なリソソームの輸送や,リソソームとほかの膜オルガネラ(後期エンドソームやファゴソームなど)との融合に関与する(図4).多細胞生物種間で高度に保存されており,ショウジョウバエや線虫ではARL8は1種類であるが,ヒトやマウスでは非常に相同性の高いARL8AとARL8Bが存在する46).
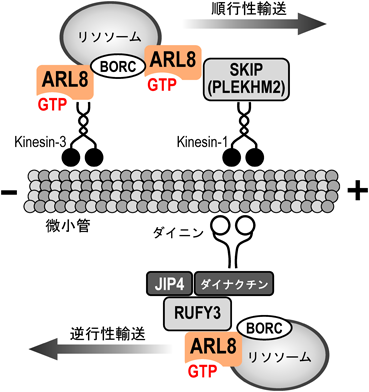
ARL8は,タンパク質複合体BORCの働きにより,リソソームに局在すると考えられている.GTP結合型ARL8は,キネシンアダプターであるSKIP(PLEKHM2)との相互作用を介してkinesin-1を,あるいは直接kinesin-3と相互作用することで,それらのモータータンパク質をリソソームにリクルートし,微小管プラス端へのリソソーム輸送に寄与する.また,GTP結合型ARL8は,RUFY3との相互作用を介してJIP4-ダイニン-ダイナクチン複合体をリソソームにリクルートすることで,微小管マイナス端への輸送にも関与すると考えられている.
ARL8のリソソーム局在化には,BORCと呼ばれるタンパク質複合体が関与すると考えられている47).リソソーム膜に局在するARL8は,エフェクターであるSKIP(PLEKHM2)との相互作用を介して,kinesin-1をリソソームにリクルートし,微小管プラス端方向へのリソソームの輸送(順行性輸送)に寄与している48).キネシンアダプターとしてのSKIPの機能は自己会合(N末端領域とC末端領域の結合)により抑制されているが,ARL8がSKIPのN末端領域のRUNドメインと結合すると,その自己会合が解除され,SKIPはkinesin-1と結合できるようになると考えられている49).また,ARL8はkinesin-3と直接相互作用し,リソソームだけでなく,シナプス小胞前駆体の微小管プラス端方向への輸送にも寄与することが報告されている50–52).
一方,ARL8のGTP結合型は,RUFY3との相互作用を介して,JIP4-ダイニン-ダイナクチン複合体をリソソームにリクルートし,リソソームの逆行性輸送にも寄与する可能性が指摘されている53, 54).RUFY3はRUNドメインと二つのcoiled-coil領域を有するタンパク質であるが,前述のSKIPとは異なり,RUNドメインではなくC末端寄りのcoiled-coil領域がARL8に重要だと考えられている.SKIPやRUFY3とARL8との相互作用が,どのような局面で使い分けられているかはわかっていない.
リソソームと膜オルガネラ(後期エンドソームなど)との融合は,それらの膜オルガネラに含有された物質分解に重要であるが,この融合過程には,ARL8とHOPS複合体との相互作用が重要だと考えられている.HOPS複合体は,リソソームの融合過程において係留因子として働くタンパク質複合体であり,その構成因子であるVPS41がARL8と相互作用することが報告されている55–57).また最近では,TGNから出芽したLAMP陽性小胞が後期エンドソーム・リソソームに輸送される過程に,VPS41とARL8の相互作用が関与する可能性が示唆されている58).
これまでに我々の研究グループは,個体レベルにおけるARL8の生理的役割を解析する目的で,線虫やマウスを用いた研究を進めてきた.線虫でのARL8欠損体は母性胚性致死であり,マクロファージ様細胞であるcoelomocyteにおいては,細胞外から取り込まれた物質が後期エンドソームまで輸送されるが,後期エンドソームとリソソームの融合が正常に進行しない59).また,アポトーシス細胞を含むファゴソームとリソソームの融合も不全であり,アポトーシス細胞のクリアランスが低下する55).したがって,これらの細胞においてARL8は,細胞外から取り込まれた物質のリソソームでの分解に重要だと考えられる.
マウスにおいては,ARL8は非常に相同性の高いArl8aとArl8bが存在するが,全身性Arl8b欠損マウス(以下,Arl8b欠損マウス)の大部分は胎生致死あるいは出生直後に致死となる.またArl8b欠損マウスの胎仔は,体長の縮小や脳形成の不全といった胚発生の異常を示す.マウス胚発生において,胎盤形成前の胎仔の発育には,卵黄嚢を介した母体由来成分の胎仔への供給が重要である.たとえば,母体由来のタンパク質(アルブミンなど)は,卵黄嚢内胚葉(visceral yolk sac endoderm)でエンドサイトーシスにより取り込まれた後,リソソームで分解され,それらの分解物が栄養素として胎仔に供給されることが正常な胚発生に重要である.興味深いことに,Arl8b欠損マウスの卵黄嚢内胚葉では,エンドサイトーシスされた物質のリソソームへの輸送が異常となり,母体由来のアルブミンやIgGが多数の小胞状構造に蓄積していた.また,卵黄嚢内胚葉特異的Arl8b欠損マウスにおいても,胎仔の体長縮小や卵黄嚢内胚葉における母体由来タンパク質の蓄積が観察された.そこで,卵黄嚢内胚葉特異的Arl8b欠損マウスの胎仔組織について,各種アミノ酸含有量を測定したところ,必須アミノ酸を含むさまざまなアミノ酸量が低下していた.以上により,Arl8bは卵黄嚢内胚葉において母体由来タンパク質のリソソームでの分解に重要であり,卵黄嚢内胚葉におけるArl8bの機能欠損は胎仔への栄養素供給低下を引き起こし,発育不良となることが示唆された60)(図5).
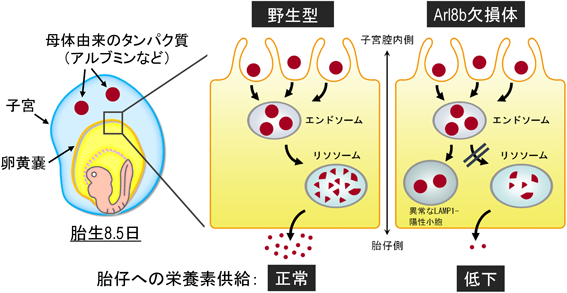
胎盤形成以前のマウス初期胚(胎生8.5日)では,母体由来のタンパク質などは,エンドサイトーシスにより卵黄嚢内胚葉に取り込まれた後,リソソームで分解され,それらの分解物は栄養素として胎仔へと供給される.Arl8b欠損マウスでは,卵黄嚢内胚葉におけるリソソーム分解が異常となり,胎仔への栄養素供給が低下して,成長障害などを引き起こす.(文献60より改変)
一方,卵黄嚢内胚葉特異的Arl8b欠損マウスにおいては,Arl8b欠損マウスでみられた脳形成の異常は観察されないことから,胎仔におけるArl8bの発現も正常な胚発生に重要であると考えられた.我々はArl8b欠損マウスの形態学的および遺伝子発現解析から,Arl8bが脳背側正中線の運命決定に重要であることを見いだした.脳背側正中線を構成する細胞は,BMP(bone morphogenetic protein)シグナルによって運命決定される細胞群であるが,Arl8b欠損マウスでは,この細胞群でBMPシグナルが異常に増強していた.この原因についての詳細は不明であるが,BMPリガンドで刺激を受けたBMP受容体はエンドサイトーシス後にリソソームで分解され,BMPシグナルのOFFに寄与すると考えられていることから,Arl8b欠損によりBMP受容体の分解不全が生じ,シグナルが持続的に活性化されることで,背側正中線の運命決定が異常となる可能性が考えられた61).
全身性Arl8b欠損マウスの一部は成体となるが,そのような個体を用いた解析から,Arl8bが形質細胞様樹状細胞(plasmacytoid dendritic cells,以下pDC)においてTLR7を介したI型インターフェロン産生に必要であることも示されている.TLR7はウイルスRNAを認識するパターン認識受容体であり,pDCにおいて主にリソソームに局在し,リガンドの結合により活性化されて,I型インターフェロン産生応答を惹起する.この細胞応答には,リガンド刺激に応答してTLR7陽性のリソソームが微小管依存的に細胞辺縁部への移行することが重要であり,Arl8bはエフェクターであるSKIP(PLEKHM2)との相互作用を介して,キネシンをTLR7陽性リソソームにリクルートすることで,その細胞辺縁部への移行を促進し,I型インターフェロン産生に寄与すると考えられている62, 63).
結腸腺がんにおいて,ARL9の遺伝子発現が上昇しており,その高発現が患者の予後不良と関連していることが報告されている64).細胞内局在や機能についてはわかっていない.
緑色蛍光タンパク質を結合したARL10(ARL10-GFP)がTOM20とよく共局在することから,主にミトコンドリアに局在すると考えられている.一次構造上の特徴として,そのN末端領域には,他のARF/ARLにはみられない膜貫通ドメインが存在する.ARL10のミトコンドリアへの局在化には,その膜貫通ドメインおよび隣接する塩基性アミノ酸に富む領域が関与すると考えられている65).
がん抑制遺伝子の候補として同定されており[別名ADP-ribosylation factor-like tumor suppressor gene 1(ARLTS1)]66),骨髄性悪性腫瘍や前立腺がんとの関連性が指摘されている.マクロファージで多く発現しており,リポ多糖(LPS)刺激によりARL11の発現が上昇する.また,LPS刺激に伴うERK1/2やp38のリン酸化,および炎症性サイトカインの産生が,ARL11の発現抑制により低下することや,ARL11とリン酸化ERKが共免疫沈降することが報告されている67).細胞内局在に関しては,ARL11-GFPの発現細胞を用いた解析から,細胞質と核内をシャトルすることが報告されている.また,動脈硬化のモデルマウス(ApoEノックアウトマウス)において,ARL11の発現が上昇すること,およびARL11の発現抑制により,マウスの大動脈における脂質沈着とアテローム性プラークの形成が減少し,炎症性サイトカインのレベルやマクロファージのM1型への極性化が低下することが報告されている68).この報告では,ARL11がJAK2と相互作用し,その安定性を介して,JAK2/STAT1経路に関与することが示唆されている.
ARL13Aについては,精巣での発現を示すデータベース情報があるが,機能的な解析は行われていない.
ARL13Bは運動性および非運動性繊毛の形成・機能に関与している.他のARF/ARLファミリーとは異なり,N末端領域に存在する低分子量Gタンパク質としてのドメインに加えて,約200アミノ酸からなるC末端領域を有する.そのC末端領域には,繊毛局在化に関与するRVxPモチーフや,coiled-coil領域,プロリンリッチ領域が存在する.また一般的なARFとは異なり,N末端近傍のシステイン残基へのパルミトイル化修飾が,ARL13Bの膜局在に必要である69, 70).
ARL13Bは,嚢胞腎の表現型を示すゼブラフィッシュ変異体scorpionの原因遺伝子として同定された71, 72).その変異体では前腎輸菅の繊毛形成不全が観察され,ARL13Bが繊毛形成・機能に関与することが初めて示唆された.また,種々の胚発生異常を示すマウス変異体hnnの原因遺伝子がARL13Bであることも報告され,その変異体においては,ノード繊毛の短小化や軸糸の形成異常,Shhシグナル伝達系の異常が観察されている73).さらにARL13Bは,繊毛性疾患ジュベール症候群(Joubert syndrome)の原因遺伝子の一つとしても知られている74).
興味深いことにARL13Bは,BARTと呼ばれるタンパク質と協調して,前述のARL3に対するGEF活性を有することが報告されている14–16).これらの研究では,精製タンパク質を用いたGEF活性の検討に加えて,培養細胞(HEK293T, IMCD3)にARL13Bを過剰発現することで,内在性ARL3のGTP結合型が増加することが示されている.ジュベール症候群でみられる遺伝子変異を有するARL13Bでは,ARL3に対するGEF活性を示さないことから,ARL13BのARL3-GEF活性喪失と疾患発症との関連性が指摘されている.また,ARL13Bのグアニンヌクレオチドに対する親和性が一般的なARFタンパク質に比べて低いことや,ウシ脳抽出液中にARL13Bに対するGAP活性が存在することが報告されている.
ARL13Bの相互作用因子としては,INPP5E(inositol polyphosphate-5-phosphatase E)が知られている.INPP5Eはジュベール症候群の原因遺伝子の一つであり75),そのC末端がプレニル化修飾を受けることが知られている.INPP5Eは前述のPDE6Dと結合して繊毛内に移行し,ARL3の作用でPDE6Dが解離した後,ARL13Bと結合することで,繊毛内に持続的に存在するようになると考えられている76, 77).
ARL13Bの繊毛局在性にはC末端付近に存在するRVxPモチーフ(ヒトではRVEP)が重要であることが知られている78–80).近年,GDP結合型のRAB8とTransportin1(TNPO1)が協調的に,ARL13BのRVxP motifを含むC末端領域に結合し,ARL13Bの繊毛局在化に寄与することが報告されている81)(図6).GTP結合型のRAB8では,RAB8-TNPO1-ARL13Bの三者複合体は形成されないことから,以下のようなモデルが想定されている.1)パルミトイル化修飾を受けたARL13Bは細胞膜に局在し,RVxPモチーフ依存的にGDP結合型RAB8とTNPO1の両者に結合し,三者複合体が形成される.2)三者複合体はTNPO1の作用により,Transition zone(一次繊毛の基部に存在する構造体で,繊毛局在性タンパク質の出入りを制御する)の障壁を越え,側方拡散により細胞膜から一次繊毛の膜に移行する.3)一次繊毛内において,RAB8に対するGEF(Rabin8やRPGR)の作用によりRAB8はGTP型となり,三者複合体は解離して,ARL13B単体が一次繊毛の膜に局在するようになる.このような一次繊毛–細胞質間の物質輸送におけるRAB8の役割と,核–細胞質間の物質輸送における低分子量Gタンパク質RANの役割の類似性が指摘されている81).また,Tubbyドメインを有するTULP3も,ARL13Bの繊毛局在性に関与することが示唆されている82).
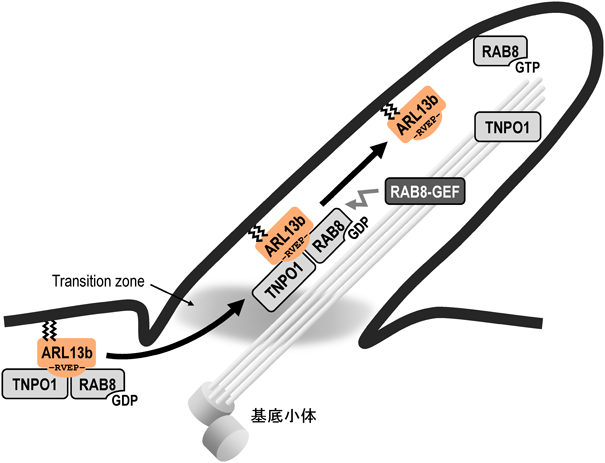
ARL13BのC末端付近には,繊毛局在化に関与するRVxPモチーフ(ヒトではRVEP)が存在する.細胞質においてGDP結合型RAB8とTransportin1(TNPO1)が,RVxPモチーフ依存的にARL13Bに結合し,それらの三者複合体は繊毛内に輸送される.繊毛内ではRabin8やRPGR(RAB8のGEF)の作用によりRAB8はGTP結合型となり,三者は解離し,遊離したARL13Bは一次繊毛に局在するようになる.
非小細胞肺がんの組織において,ARL14の発現が正常組織と比較して高いことが報告されており83),肺腺がん細胞でARL14をノックダウンすると,ERK/p38シグナル経路の減弱と細胞増殖抑制が観察されることが報告されている84).また,ARL14遺伝子の転写調節に関わる領域の一塩基多型(SNP)と,重症マラリアの発症リスクとの関連性も指摘されている85).細胞内におけるARL14の役割は不明な点が多いが,RNA干渉(RNAi)を用いたスクリーニングから,ARL14は樹状細胞におけるMHCクラスIIの細胞内輸送に関与することが示唆されている.ARL14はC11ORF46というタンパク質との相互作用を介して,MHCクラスII陽性小胞にミオシンモーターをリクルートし,アクチン繊維依存的な細胞内輸送を制御するというモデルが提唱されている86).また,近位依存性ビオチン化法(miniTurboID)を用いた解析から,ARL14近傍因子の候補としてPLD1が同定されており,PLD1の活性制御に関与する可能性も示唆されている65).ARF14-GFPを用いた解析から,ARL14は細胞膜や細胞内の小胞状構造(一部は初期エンドソームマーカー陽性)に局在すると考えられている.また,ARL14のN末端の両親媒性αヘリックス構造とGFPの融合タンパク質がARL14-GFPと同様の細胞内局在を示すことから,そのαヘリックス構造がARL14の細胞内局在を規定すると考えられている87).
各種のGWAS(genome-wide association study)解析から,種々の代謝性疾患との関連性が指摘されている88–92).ARL15のN末端から2番目のアミノ酸はセリン残基(一般的なARFではグリシン残基)であり,ミリストイル化修飾は受けない.一方,ARL15のN末端近傍には生物種間でよく保存されたシステイン残基が複数存在し,それらがパルミトイル化修飾されることで,細胞膜やTGNに存在すると考えられている93).ARL15の相互作用因子としては,Mg2+トランスポーターであるCNNMタンパク質や,イオンチャネルであるTRPM7が報告されており,細胞内のマグネシウム濃度調節に関与することが示唆されている.
ARL15とCNNMの構造化学的解析から,ARL15とCNNM2のCBSドメインが相互作用することが示されている94, 95).ARL15はCNNMやTRPM7と相互作用することで,それらを介した金属イオンの輸送調節に関与する可能性がある95, 96).また,ARL15を過剰発現あるいは欠損させると,CNNM3の糖鎖修飾が変化することから,CNNMの細胞内輸送への関与が示唆されている94).また,ARL15の大腸菌リコンビナントタンパク質を用いた解析から,ARL15のGTPに対する親和性は,一般的な低分子量Gタンパク質に比べて非常に低い(約20 µM程度)ことが報告されている95).
ARL15ノックアウトマウスは出生後離乳前に致死であり,口蓋形成不全の表現型を示し,ARL15ノックアウトマウス由来のMEF(mouse embryonic fibroblast)では,野生型に比べて細胞移動能が低下することが報告されている97).一方,ARL15ヘテロ欠損マウスを用いた解析から,成体における褐色脂肪組織や骨形成へのARL15の関与が示唆されている.
繊毛形成がみられる真核生物種でよく保存されており,一次繊毛の形成・機能に関与すると考えられている.ARL16欠損細胞では,IFT140およびINPP5Eの繊毛局在性が低下し,それらがゴルジ体に蓄積することから,ARL16がゴルジ体–一次繊毛間輸送に関与することが示唆されている98).また,ARFのGAPとして知られるELMOD1/3との機能的関連性が指摘されている99).
一方,RIG-Iを介するシグナル伝達系にARL16が関与する可能性も報告されている.RIG-IはRNAウイルス由来の核酸を認識し,インターフェロンを誘導することで宿主の抗ウイルス応答を活性化するセンサーであるが,ARL16のGTP結合型がRIG-IのC末端ドメイン(RNA認識領域)に結合し,RIG-IとRNAの結合を阻害することで,RIG-Iの活性化を阻害することが報告されている100).
以上,ARL分子群に関する知見を列挙させていただいたが,ARFに比べると非常に多様な細胞機能に関与することがおわかりいただけたかと思う.しかし,それらの制御機構に関しては,いまだにほとんどわかっていない.たとえば大部分のARLについては,GEFやGAPは未同定であり,細胞内においてARLの活性(グアニンヌクレオチド結合状態)がどのように調節されているかは未解明である.ARLの生化学的特性に関しては,ARL13BやARL15にみられるように,グアニンヌクレオチドに対する親和性が非常に低いものも存在することから,ARLの活性制御は一般的なGタンパク質とはかなり異なる可能性もある.今後,細胞レベルの解析に加え,精製標品などを用いた生化学的解析なども,ARLの素性を明らかにする上で必要であろう.また,個体レベルにおけるARLの生理的役割に関しても多くは不明であり,それらの解明も待たれる.ARL分子群に関する今後のさらなる解析が,さまざまな細胞応答の調節機構の理解と,低分子量Gタンパク質研究の発展へとつながることを期待したい.
本稿で御紹介させていただいたARL8に関する個体レベルでの機能解析は,多くの共同研究者の方々とともに行われたものであり,この場を借りて関係者の皆様に深く御礼申し上げます.
1) Sztul, E., Chen, P.-W., Casanova, J.E., Cherfils, J., Dacks, J.B., Lambright, D.G., Lee, F.-J.S., Randazzo, P.A., Santy, L.C., Schürmann, A., et al. (2019) ARF GTPases and their GEFs and GAPs: Concepts and challenges. Mol. Biol. Cell, 30, 1249–1271.
2) Yu, C.-J. & Lee, F.-J.S. (2017) Multiple activities of Arl1 GTPase in the trans-Golgi network. J. Cell Sci., 130, 1691–1699.
3) Valkenburgh, H., Shern, J.F., Sharer, D.J., Zhu, X., & Kahn, R.A. (2001). J. Biol. Chem., 276, 22826–22837.
4) Lu, L. & Hong, W. (2003) Interaction of Arl1-GTP with GRIP domains recruits autoantigens Golgin-97 and Golgin-245/p230 onto the Golgi. Mol. Biol. Cell, 14, 3767–3781.
5) Ishida, M. & Bonifacino, J.S. (2019) ARFRP1 functions upstream of ARL1 and ARL5 to coordinate recruitment of distinct tethering factors to the trans-Golgi network. J. Cell Biol., 218, 3681–3696.
6) D’Souza, R.S., Semus, R., Billings, E.A., Meyer, C.B., Conger, K., & Casanova, J.E. (2014) Rab4 orchestrates a small GTPase cascade for recruitment of adaptor proteins to early endosomes. Curr. Biol., 24, 1187–1198.
7) Francis, J.W., Newman, L.E., Cunningham, L.A., & Kahn, R.A. (2017) A trimer consisting of the tubulin-specific chaperone D (TBCD), regulatory GTPase ARL2, and β-tubulin is required for maintaining the microtubule network. J. Biol. Chem., 292, 4336–4349.
8) Newman, L.E., Zhou, C., Mudigonda, S., Mattheyses, A.L., Paradies, E., Marobbio, C.M.T., & Kahn, R.A. (2014) The ARL2 GTPase is required for mitochondrial morphology, motility, and maintenance of ATP levels. PLoS One, 9, e99270.
9) Newman, L.E., Schiavon, C.R., Turn, R.E., & Kahn, R.A. (2017) The ARL2 GTPase regulates mitochondrial fusion from the intermembrane space. Cell. Logist., 7, e1340104.
10) Zhou, C., Cunningham, L., Marcus, A.I., Li, Y., & Kahn, R.A. (2006) Arl2 and Arl3 regulate different microtubule-dependent processes. Mol. Biol. Cell, 17, 2476–2487.
11) Ma, D., Lin, K.-Y., Suresh, D., Lin, J., Gujar, M.R., Aung, H.Y., Tan, Y.S., Gao, Y., Vincent, A.S., Chen, T., et al. (2024) Arl2 GTPase associates with the centrosomal protein Cdk5rap2 to regulate cortical development via microtubule organization. PLoS Biol., 22, e3002751.
12) Powell, L., Samarakoon, Y.H., Ismail, S., & Sayer, J.A. (2021) ARL3, a small GTPase with a functionally conserved role in primary cilia and immune synapses. Small GTPases, 12, 167–176.
13) Fansa, E.K. & Wittinghofer, A. (2016) Sorting of lipidated cargo by the Arl2/Arl3 system. Small GTPases, 7, 222–230.
14) ElMaghloob, Y., Sot, B., McIlwraith, M.J., Garcia, E., Yelland, T., & Ismail, S. (2021) ARL3 activation requires the co-GEF BART and effector-mediated turnover. eLife, 10, e64624.
15) Gotthardt, K., Lokaj, M., Koerner, C., Falk, N., Gießl, A., & Wittinghofer, A. (2015) A G-protein activation cascade from Arl13B to Arl3 and implications for ciliary targeting of lipidated proteins. eLife, 4, e11859.
16) Ivanova, A.A., Caspary, T., Seyfried, N.T., Duong, D.M., West, A.B., Liu, Z., & Kahn, R.A. (2017) Biochemical characterization of purified mammalian ARL13B protein indicates that it is an atypical GTPase and ARL3 guanine nucleotide exchange factor (GEF). J. Biol. Chem., 292, 11091–11108.
17) Veltel, S., Gasper, R., Eisenacher, E., & Wittinghofer, A. (2008) The retinitis pigmentosa 2 gene product is a GTPase-activating protein for Arf-like 3. Nat. Struct. Mol. Biol., 15, 373–380.
18) Wright, K.J., Baye, L.M., Olivier-Mason, A., Mukhopadhyay, S., Sang, L., Kwong, M., Wang, W., Pretorius, P.R., Sheffield, V.C., Sengupta, P., et al. (2011) An ARL3-UNC119-RP2 GTPase cycle targets myristoylated NPHP3 to the primary cilium. Genes Dev., 25, 2347–2360.
19) Veltel, S., Kravchenko, A., Ismail, S., & Wittinghofer, A. (2008) Specificity of Arl2/Arl3 signaling is mediated by a ternary Arl3-effector-GAP complex. FEBS Lett., 582, 2501–2507.
20) Patel, M., Chiang, T.-C., Tran, V., Lee, F.-J.S., & Côté, J.-F. (2011) The Arf family GTPase Arl4A complexes with ELMO proteins to promote actin cytoskeleton remodeling and reveals a versatile Ras-binding domain in the ELMO proteins family. J. Biol. Chem., 286, 38969–38979.
21) Chiang, T.-S., Lin, M.-C., Tsai, M.-C., Chen, C.-H., Jang, L.-T., & Lee, F.-J.S. (2019) ADP-ribosylation factor-like 4A interacts with Robo1 to promote cell migration by regulating Cdc42 activation. Mol. Biol. Cell, 30, 69–81.
22) Lin, Y.-C., Chiang, T.-C., Liu, Y.-T., Tsai, Y.-T., Jang, L.-T., & Lee, F.-J.S. (2011) ARL4A acts with GCC185 to modulate Golgi complex organization. J. Cell Sci., 124, 4014–4026.
23) Lin, S.-J., Lin, M.-C., Liu, T.-J., Tsai, Y.-T., Tsai, M.-T., & Lee, F.-J.S. (2023) Endosomal Arl4A attenuates EGFR degradation by binding to the ESCRT-II component VPS36. Nat. Commun., 14, 7859.
24) Hofmann, I., Thompson, A., Sanderson, C.M., & Munro, S. (2007) The Arl4 family of small G proteins can recruit the cytohesin Arf6 exchange factors to the plasma membrane. Curr. Biol., 17, 711–716.
25) Chiang, T.-S., Wu, H.-F., & Lee, F.-J.S. (2017) ADP-ribosylation factor-like 4C binding to filamin-A modulates filopodium formation and cell migration. Mol. Biol. Cell, 28, 3013–3028.
26) Harada, A., Matsumoto, S., Yasumizu, Y., Akama, T., Eguchi, H., & Kikuchi, A. (2021) Localization of KRAS downstream target ARL4C to invasive pseudopods accelerates pancreatic cancer cell invasion. eLife, 10, e66721.
27) Matsumoto, S., Fujii, S., & Kikuchi, A. (2017) Arl4c is a key regulator of tubulogenesis and tumourigenesis as a target gene of Wnt-β-catenin and growth factor-Ras signalling. J. Biochem., 161, 27–35.
28) Fujii, S., Matsumoto, S., Nojima, S., Morii, E., & Kikuchi, A. (2015) Arl4c expression in colorectal and lung cancers promotes tumorigenesis and may represent a novel therapeutic target. Oncogene, 34, 4834–4844.
29) Zhang, J., Zhang, Q., Sun, C., Huang, Y., Zhang, J., & Wang, Q. (2020) Clinical relevance of ARF/ARL family genes and oncogenic function of ARL4C in endometrial cancer. Biomed. Pharmacother., 125, 110000.
30) Kanai, R., Uehara, T., Yoshizawa, T., Kamakura, M., Nakajima, T., Kinugawa, Y., Iwaya, M., Asaka, S., Kitazawa, M., Nagaya, T., et al. (2023) ARL4C is associated with epithelial-to-mesenchymal transition in colorectal cancer. BMC Cancer, 23, 478.
31) Fujii, S., Ishibashi, T., Kokura, M., Fujimoto, T., Matsumoto, S., Shidara, S., Kurppa, K.J., Pape, J., Caton, J., Morgan, P.R., et al. (2022) RAF1-MEK/ERK pathway-dependent ARL4C expression promotes ameloblastoma cell proliferation and osteoclast formation. J. Pathol., 256, 119–133.
32) Li, C.-C., Chiang, T.-C., Wu, T.-S., Pacheco-Rodriguez, G., Moss, J., & Lee, F.-J.S. (2007) ARL4D recruits cytohesin-2/ARNO to modulate actin remodeling. Mol. Biol. Cell, 18, 4420–4437.
33) Yamauchi, J., Miyamoto, Y., Torii, T., Mizutani, R., Nakamura, K., Sanbe, A., Koide, H., Kusakawa, S., & Tanoue, A. (2009) Valproic acid-inducible Arl4D and cytohesin-2/ARNO, acting through the downstream Arf6, regulate neurite outgrowth in N1E-115 cells. Exp. Cell Res., 315, 2043–2052.
34) Lin, S.-J., Huang, C.-F., Wu, T.-S., Li, C.-C., & Lee, F.-J.S. (2020) Arl4D-EB1 interaction promotes centrosomal recruitment of EB1 and microtubule growth. Mol. Biol. Cell, 31, 2348–2362.
35) Houghton, F.J., Bellingham, S.A., Hill, A.F., Bourges, D., Ang, D.K., Gemetzis, T., Gasnereau, I., & Gleeson, P.A. (2012) Arl5b is a Golgi-localised small G protein involved in the regulation of retrograde transport. Exp. Cell Res., 318, 464–477.
36) Rosa-Ferreira, C., Christis, C., Torres, I.L., & Munro, S. (2015) The small G protein Arl5 contributes to endosome-to-Golgi traffic by aiding the recruitment of the GARP complex to the Golgi. Biol. Open, 4, 474–481.
37) Houghton, F.J., Makhoul, C., Cho, E.H., Williamson, N.A., & Gleeson, P.A. (2022) Interacting partners of Golgi-localized small G protein Arl5b identified by a combination of in vivo proximity labelling and GFP-Trap pull down. FEBS Lett., 596, 2382–2399.
38) Jaimon, E., Tripathi, A., Khurana, A., Ghosh, D., Sugatha, J., & Datta, S. (2021) Binding with heat shock cognate protein HSC70 fine-tunes the Golgi association of the small GTPase ARL5B. J. Biol. Chem., 297, 101422.
39) Nachury, M.V. (2018) The molecular machines that traffic signaling receptors into and out of cilia. Curr. Opin. Cell Biol., 51, 124–131.
40) Nakayama, K. & Katoh, Y. (2018) Ciliary protein trafficking mediated by IFT and BBSome complexes with the aid of kinesin-2 and dynein-2 motors. J. Biochem., 163, 155–164.
41) Jin, H., White, S.R., Shida, T., Schulz, S., Aguiar, M., Gygi, S.P., Bazan, J.F., & Nachury, M.V. (2010) The conserved Bardet-Biedl syndrome proteins assemble a coat that traffics membrane proteins to cilia. Cell, 141, 1208–1219.
42) Xue, B., Liu, Y.-X., Dong, B., Wingfield, J.L., Wu, M., Sun, J., Lechtreck, K.F., & Fan, Z.-C. (2020) Intraflagellar transport protein RABL5/IFT22 recruits the BBSome to the basal body through the GTPase ARL6/BBS3. Proc. Natl. Acad. Sci. USA, 117, 2496–2505.
43) Kobayashi, T., Hori, Y., Ueda, N., Kajiho, H., Muraoka, S., Shima, F., Kataoka, T., Kontani, K., & Katada, T. (2009) Biochemical characterization of missense mutations in the Arf/Arl-family small GTPase Arl6 causing Bardet-Biedl syndrome. Biochem. Biophys. Res. Commun., 381, 439–442.
44) Liew, G.M., Ye, F., Nager, A.R., Murphy, J.P., Lee, J.S., Aguiar, M., Breslow, D.K., Gygi, S.P., & Nachury, M.V. (2014) The intraflagellar transport protein IFT27 promotes BBSome exit from cilia through the GTPase ARL6/BBS3. Dev. Cell, 31, 265–278.
45) Liu, Y., Zhang, R., & Fan, Z. (2023) RABL4/IFT27 in a nucleotide-independent manner promotes phospholipase D ciliary retrieval via facilitating BBSome reassembly at the ciliary tip. J. Cell. Physiol., 238, 549–565.
46) Hofmann, I. & Munro, S. (2006) An N-terminally acetylated Arf-like GTPase is localised to lysosomes and affects their motility. J. Cell Sci., 119, 1494–1503.
47) Pu, J., Schindler, C., Jia, R., Jarnik, M., Backlund, P., & Bonifacino, J.S. (2015) BORC, a multisubunit complex that regulates lysosome positioning. Dev. Cell, 33, 176–188.
48) Rosa-Ferreira, C. & Munro, S. (2011) Arl8 and SKIP act together to link lysosomes to kinesin-1. Dev. Cell, 21, 1171–1178.
49) Keren-Kaplan, T. & Bonifacino, J.S. (2021) ARL8 relieves SKIP autoinhibition to enable coupling of lysosomes to kinesin-1. Curr. Biol., 31, 540–554.
50) Wu, Y.E., Huo, L., Maeder, C.I., Feng, W., & Shen, K. (2013) The balance between capture and dissociation of presynaptic proteins controls the spatial distribution of synapses. Neuron, 78, 994–1011.
51) Niwa, S., Lipton, D.M., Morikawa, M., Zhao, C., Hirokawa, N., Lu, H., & Shen, K. (2016) Autoinhibition of a neuronal kinesin UNC-104/KIF1A regulates the size and density of synapses. Cell Rep., 16, 2129–2141.
52) Guardia, C.M., Farías, G.G., Jia, R., Pu, J., & Bonifacino, J.S. (2016) BORC Functions upstream of kinesins 1 and 3 to coordinate Regional movement of lysosomes along different microtubule tracks. Cell Rep., 17, 1950–1961.
53) Keren-Kaplan, T., Sarić, A., Ghosh, S., Williamson, C.D., Jia, R., Li, Y., & Bonifacino, J.S. (2022) RUFY3 and RUFY4 are ARL8 effectors that promote coupling of endolysosomes to dynein-dynactin. Nat. Commun., 13, 1506.
54) Kumar, G., Chawla, P., Dhiman, N., Chadha, S., Sharma, S., Sethi, K., Sharma, M., & Tuli, A. (2022) RUFY3 links Arl8b and JIP4-Dynein complex to regulate lysosome size and positioning. Nat. Commun., 13, 1540.
55) Sasaki, A., Nakae, I., Nagasawa, M., Hashimoto, K., Abe, F., Saito, K., Fukuyama, M., Gengyo-Ando, K., Mitani, S., Katada, T., et al. (2013) Arl8/ARL-8 functions in apoptotic cell removal by mediating phagolysosome formation in Caenorhabditis elegans. Mol. Biol. Cell, 24, 1584–1592.
56) Khatter, D., Raina, V.B., Dwivedi, D., Sindhwani, A., Bahl, S., & Sharma, M. (2015) The small GTPase Arl8b regulates assembly of the mammalian HOPS complex on lysosomes. J. Cell Sci., 128, 1746–1761.
57) Garg, S., Sharma, M., Ung, C., Tuli, A., Barral, D.C., Hava, D.L., Veerapen, N., Besra, G.S., Hacohen, N., & Brenner, M.B. (2011) Lysosomal trafficking, antigen presentation, and microbial killing are controlled by the Arf-like GTPase Arl8b. Immunity, 35, 182–193.
58) Sanzà, P., van der Beek, J., Draper, D., de Heus, C., Veenendaal, T., ten Brink, C., Farías, G.G., Liv, N., & Klumperman, J. (2025) VPS41 recruits biosynthetic LAMP-positive vesicles through interaction with Arl8b. J. Cell Biol., 224, e202405002.
59) Nakae, I., Fujino, T., Kobayashi, T., Sasaki, A., Kikko, Y., Fukuyama, M., Gengyo-Ando, K., Mitani, S., Kontani, K., & Katada, T. (2010) The arf-like GTPase Arl8 mediates delivery of endocytosed macromolecules to lysosomes in Caenorhabditis elegans. Mol. Biol. Cell, 21, 2434–2442.
60) Oka, M., Hashimoto, K., Yamaguchi, Y., Saitoh, S., Sugiura, Y., Motoi, Y., Honda, K., Kikko, Y., Ohata, S., Suematsu, M., et al. (2017) Arl8b is required for lysosomal degradation of maternal proteins in the visceral yolk sac endoderm of mouse embryos. J. Cell Sci., 130, 3568–3577.
61) Hashimoto, K., Yamaguchi, Y., Kishi, Y., Kikko, Y., Takasaki, K., Maeda, Y., Matsumoto, Y., Oka, M., Miura, M., Ohata, S., et al. (2019) Loss of the small GTPase Arl8b results in abnormal development of the roof plate in mouse embryos. Genes Cells, 24, 436–448.
62) Saitoh, S.-I., Abe, F., Kanno, A., Tanimura, N., Saitoh, Y.M., Fukui, R., Shibata, T., Sato, K., Ichinohe, T., Hayashi, M., et al. (2017) TLR7 mediated viral recognition results in focal type I interferon secretion by dendritic cells. Nat. Commun., 8, 1592.
63) Saitoh, S.-I., Saitoh, Y.M., Kontani, K., Sato, K., & Miyake, K. (2019) ADP-ribosylation factor-like 8b is required for the development of mouse models of systemic lupus erythematosus. Int. Immunol., 31, 225–237.
64) Yang, H., Jin, X., Gu, B., Yu-Zhang, Li, D., & Yan, L.L. (2023) ARL9 is upregulated and serves as a biomarker for a poor prognosis in colon adenocarcinoma. BMC Gastroenterol., 23, 48.
65) Quirion, L., Robert, A., Boulais, J., Huang, S., Astrain, G.B., Strakhova, R., Jo, C.H., Kherdjemil, Y., Faubert, D., Thibault, M.-P., et al. (2024) Mapping the global interactome of the ARF family reveals spatial organization in cellular signaling pathways. J. Cell Sci., 137, jcs262140.
66) Sellick, G.S., Catovsky, D., & Houlston, R.S. (2006) Familial cancer associated with a polymorphism in ARLTS1. N. Engl. J. Med., 354, 1204–1205., author reply, 1204–1205.
67) Arya, S.B., Kumar, G., Kaur, H., Kaur, A., & Tuli, A. (2018) ARL11 regulates lipopolysaccharide-stimulated macrophage activation by promoting mitogen-activated protein kinase (MAPK) signaling. J. Biol. Chem., 293, 9892–9909.
68) Zhen, Y., Yang, J., Song, J., Xing, Z., & Zheng, J. (2024) Silencing ARL11 relieved atherosclerotic inflammation and lipid deposition via retraining JAK2/STAT1 pathway. Atherosclerosis, 398, 118564.
69) Cevik, S., Hori, Y., Kaplan, O.I., Kida, K., Toivenon, T., Foley-Fisher, C., Cottell, D., Katada, T., Kontani, K., & Blacque, O.E. (2010) Joubert syndrome Arl13b functions at ciliary membranes and stabilizes protein transport in Caenorhabditis elegans. J. Cell Biol., 188, 953–969.
70) Roy, K., Jerman, S., Jozsef, L., McNamara, T., Onyekaba, G., Sun, Z., & Marin, E.P. (2017) Palmitoylation of the ciliary GTPase ARL13b is necessary for its stability and its role in cilia formation. J. Biol. Chem., 292, 17703–17717.
71) Sun, Z., Amsterdam, A., Pazour, G.J., Cole, D.G., Miller, M.S., & Hopkins, N. (2004) A genetic screen in zebrafish identifies cilia genes as a principal cause of cystic kidney. Development, 131, 4085–4093.
72) Duldulao, N.A., Lee, S., & Sun, Z. (2009) Cilia localization is essential for in vivo functions of the Joubert syndrome protein Arl13b/Scorpion. Development, 136, 4033–4042.
73) Caspary, T., Larkins, C.E., & Anderson, K.V. (2007) The graded response to Sonic Hedgehog depends on cilia architecture. Dev. Cell, 12, 767–778.
74) Cantagrel, V., Silhavy, J.L., Bielas, S.L., Swistun, D., Marsh, S.E., Bertrand, J.Y., Audollent, S., Attié-Bitach, T., Holden, K.R., Dobyns, W.B., et al. (2008) Mutations in the cilia gene ARL13B lead to the classical form of Joubert syndrome. Am. J. Hum. Genet., 83, 170–179.
75) Bielas, S.L., Silhavy, J.L., Brancati, F., Kisseleva, M.V., Al-Gazali, L., Sztriha, L., Bayoumi, R.A., Zaki, M.S., Abdel-Aleem, A., Rosti, R.O., et al. (2009) Mutations in INPP5E, encoding inositol polyphosphate-5-phosphatase E, link phosphatidyl inositol signaling to the ciliopathies. Nat. Genet., 41, 1032–1036.
76) Humbert, M.C., Weihbrecht, K., Searby, C.C., Li, Y., Pope, R.M., Sheffield, V.C., & Seo, S. (2012) ARL13B, PDE6D, and CEP164 form a functional network for INPP5E ciliary targeting. Proc. Natl. Acad. Sci. USA, 109, 19691–19696.
77) Fujisawa, S., Qiu, H., Nozaki, S., Chiba, S., Katoh, Y., & Nakayama, K. (2021) ARL3 and ARL13B GTPases participate in distinct steps of INPP5E targeting to the ciliary membrane. Biol. Open, 10, bio058843.
78) Higginbotham, H., Eom, T.-Y., Mariani, L.E., Bachleda, A., Hirt, J., Gukassyan, V., Cusack, C.L., Lai, C., Caspary, T., & Anton, E. (2012) Arl13b in primary cilia regulates the migration and placement of interneurons in the developing cerebral cortex. Dev. Cell, 23, 925–938.
79) Cevik, S., Sanders, A.A.W.M., Wijk, E.V., Boldt, K., Clarke, L., van Reeuwijk, J., Hori, Y., Horn, N., Hetterschijt, L., Wdowicz, A., et al. (2013) Active transport and diffusion barriers restrict Joubert Syndrome-associated ARL13B/ARL-13 to an Inv-like ciliary membrane subdomain. PLoS Genet., 9, e1003977.
80) Nozaki, S., Katoh, Y., Terada, M., Michisaka, S., Funabashi, T., Takahashi, S., Kontani, K., & Nakayama, K. (2016) Regulation of ciliary retrograde protein trafficking by the Joubert syndrome proteins ARL13B and INPP5E. J. Cell Sci., 130, 563–576.
81) Mahajan, D., Madugula, V., & Lu, L. (2023) Rab8 and TNPO1 are ciliary transport adaptors for GTPase Arl13b by interacting with its RVEP motif containing ciliary targeting sequence. J. Biol. Chem., 299, 104604.
82) Palicharla, V.R., Hwang, S.-H., Somatilaka, B.N., Legué, E., Shimada, I.S., Familiari, N.E., Tran, V.M., Woodruff, J.B., Liem, K.F. Jr., & Mukhopadhyay, S. (2023) Interactions between TULP3 tubby domain and ARL13B amphipathic helix promote lipidated protein transport to cilia. Mol. Biol. Cell, 34, ar18.
83) Zhang, B., Xu, A., Wu, D., Xia, W., Li, P., Wang, E., Han, R., Sun, P., Zhou, S., & Wang, R. (2021) ARL14 as a Prognostic Biomarker in Non-Small Cell Lung Cancer. J. Inflamm. Res., 14, 6557–6574.
84) Guo, F., Yuan, D., Zhang, J., Zhang, H., Wang, C., Zhu, L., Zhang, J., Pan, Y., & Shao, C. (2019) Silencing of ARL14 Gene Induces Lung Adenocarcinoma Cells to a Dormant State. Front. Cell Dev. Biol., 7, 238.
85) Adjemout, M., Gallardo, F., Torres, M., Thiam, A., Mbengue, B., Dieye, A., Marquet, S., & Rihet, P. (2024) From Genome-wide Association Studies to Functional Variants: ARL14 Cis-regulatory Variants Are Associated With Severe Malaria. J. Infect. Dis., 230, e743–e752.
86) Paul, P., van den Hoorn, T., Jongsma, M.L.M., Bakker, M.J., Hengeveld, R., Janssen, L., Cresswell, P., Egan, D.A., van Ham, M., ten Brinke, A., et al. (2011) A Genome-wide multidimensional RNAi screen reveals pathways controlling MHC class II antigen presentation. Cell, 145, 268–283.
87) Yang, F., Li, T., Peng, Z., Liu, Y., & Guo, Y. (2020) The amphipathic helices of Arfrp1 and Arl14 are sufficient to determine subcellular localizations. J. Biol. Chem., 295, 16643–16654.
88) Richards, J., Waterworth, D., O’Rahilly, S., Hivert, M.-F.F., Loos, R.J., Perry, J.R., Tanaka, T., Timpson, N.J., Semple, R.K., Soranzo, N., et al. (2009) A genome-wide association study reveals variants in ARL15 that influence adiponectin levels. PLoS Genet., 5, e1000768.
89) Mahajan, A., Go, M.J., Zhang, W., Below, J.E., Gaulton, K.J., Ferreira, T., Horikoshi, M., Johnson, A.D., Ng, M.C.Y., Prokopenko, I., et al. (2014) Genome-wide trans-ancestry meta-analysis provides insight into the genetic architecture of type 2 diabetes susceptibility. Nat. Genet., 46, 234–244.
90) Thomsen, S.K., Ceroni, A., van de Bunt, M., Burrows, C., Barrett, A., Scharfmann, R., Ebner, D., McCarthy, M.I., & Gloyn, A.L. (2016) Systematic functional characterization of candidate causal genes for type 2 diabetes risk variants. Diabetes, 65, 3805–3811.
91) Gorski, M., van der Most, P.J., Teumer, A., Chu, A.Y., Li, M., Mijatovic, V., Nolte, I.M., Cocca, M., Taliun, D., Gomez, F., et al. (2017) 1000 Genomes-based meta-analysis identifies 10 novel loci for kidney function. Sci. Rep., 7, srep45040.
92) Kashyap, S., Kumar, U., Pandey, A.K., Kanjilal, M., Chattopadhyay, P., Yadav, C., & Thelma, B.K. (2018) Functional characterisation of ADP ribosylation factor-like protein 15 in rheumatoid arthritis synovial fibroblasts. Clin. Exp. Rheumatol., 36, 581–588.
93) Wu, Y., Bai, Y., McEwan, D.G., Bentley, L., Aravani, D., & Cox, R.D. (2021) Palmitoylated small GTPase ARL15 is translocated within Golgi network during adipogenesis. Biol. Open, 10, bio058420.
94) Zolotarov, Y., Ma, C., González-Recio, I., Hardy, S., Franken, G.A., Uetani, N., Latta, F., Kostantin, E., Boulais, J., Thibault, M.-P., et al. (2021) ARL15 modulates magnesium homeostasis through N-glycosylation of CNNMs. Cell. Mol. Life Sci., 78, 5427–5445.
95) Mahbub, L., Kozlov, G., Zong, P., Lee, E.L., Tetteh, S., Nethramangalath, T., Knorn, C., Jiang, J., Shahsavan, A., Yue, L., et al. (2023) Structural insights into regulation of CNNM-TRPM7 divalent cation uptake by the small GTPase ARL15. eLife, 12, e86129.
96) Hardy, S., Zolotarov, Y., Coleman, J., Roitman, S., Khursheed, H., Aubry, I., Uetani, N., & Tremblay, M.L. (2023) PRL-1/2 phosphatases control TRPM7 magnesium-dependent function to regulate cellular bioenergetics. Proc. Natl. Acad. Sci. USA, 120, e2221083120.
97) Bai, Y., Bentley, L., Ma, C., Naveenan, N., Cleak, J., Wu, Y., Simon, M.M., Westerberg, H., Cañas, R.C., Horner, N., et al. (2023) Cleft palate and minor metabolic disturbances in a mouse global Arl15 gene knockout. FASEB J., 37, e23211.
98) Dewees, S.I., Vargová, R., Hardin, K.R., Turn, R.E., Devi, S., Linnert, J., Wolfrum, U., Caspary, T., Eliáš, M., & Kahn, R.A. (2022) Phylogenetic profiling and cellular analyses of ARL16 reveal roles in traffic of IFT140 and INPP5E. Mol. Biol. Cell, 33, ar33.
99) Turn, R.E., Hu, Y., Dewees, S.I., Devi, N., East, M.P., Hardin, K.R., Khatib, T., Linnert, J., Wolfrum, U., Lim, M.J., et al. (2022) The ARF GAPs ELMOD1 and ELMOD3 act at the Golgi and cilia to regulate ciliogenesis and ciliary protein traffic. Mol. Biol. Cell, 33, ar13.
100) Yang, Y.-K., Qu, H., Gao, D., Di, W., Chen, H.-W., Guo, X., Zhai, Z.-H., & Chen, D.-Y. (2011) ARF-like protein 16 (ARL16) inhibits RIG-I by binding with its C-terminal domain in a GTP-dependent manner. J. Biol. Chem., 286, 10568–10580.

明治薬科大学薬学部生化学研究室 教授.博士(理学).
1990年東京工業大学理学部卒業.95年同大学院生命理工学研究科バイオサイエンス専攻博士課程修了.98年東京大学薬学部助手.2002年カリフォルニア大学サンタバーバラ校博士研究員.05年東京大学薬学部助教授.15年より現職.
研究テーマと抱負低分子量Gタンパク質を切り口とした細胞応答の制御機構の理解.ARL分子群のアティピカルな生化学的性状の生理的意義も明らかにしたい.
趣味(最近あまり行けずにいるが)ライブハウスやコンサートに行くこと.

This page was created on 2025-07-15T10:58:28.204+09:00
This page was last modified on 2025-08-08T16:21:07.000+09:00
このサイトは(株)国際文献社によって運用されています。