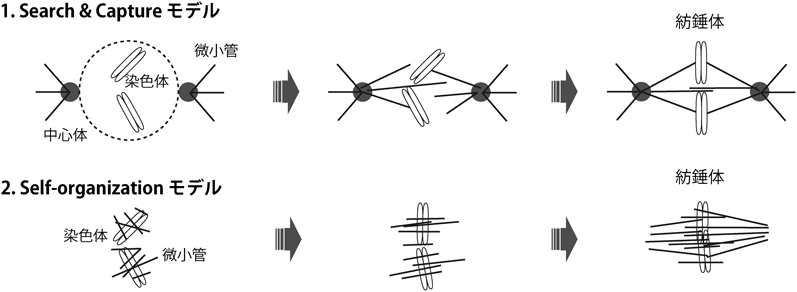
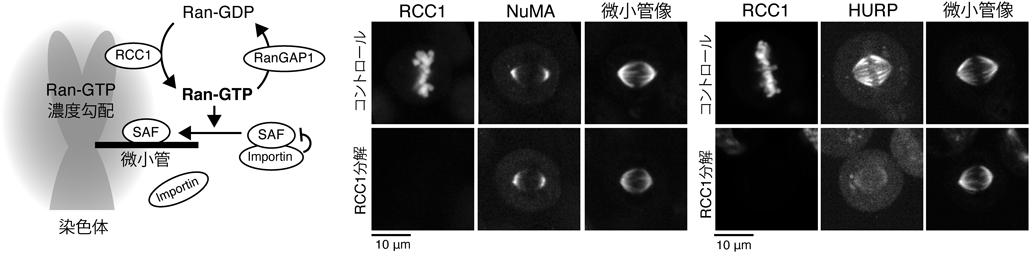
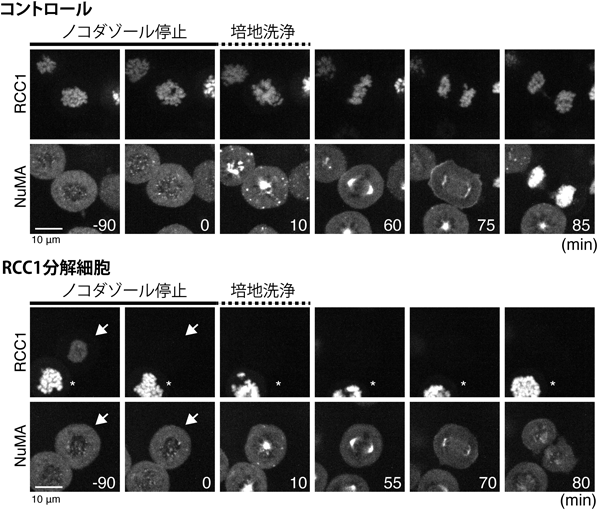
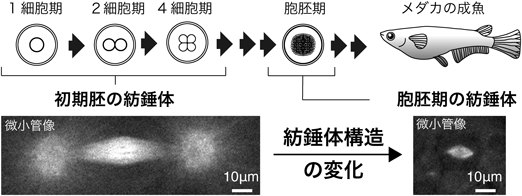
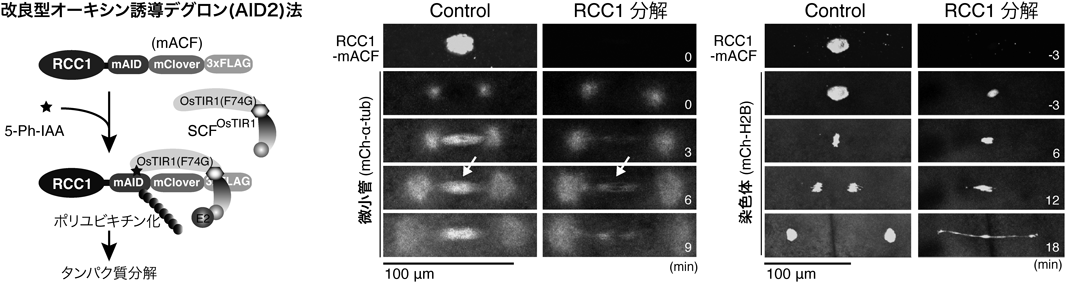
初期発生におけるRan-GTPの紡錘体形成機能とその必須性の変化Function and essentiality of Ran-based spindle assembly in early embryogenesis
沖縄科学技術大学院大学Okinawa Institute of Science and Technology Graduate University ◇ 〒904–0495 沖縄県国頭郡恩納村字谷茶1919–1 ◇ 1919–1 Tancha, Onna-son, Kunigami-gun, Okinawa 904–0495, Japan
発行日:2025年8月25日Published: August 25, 2025