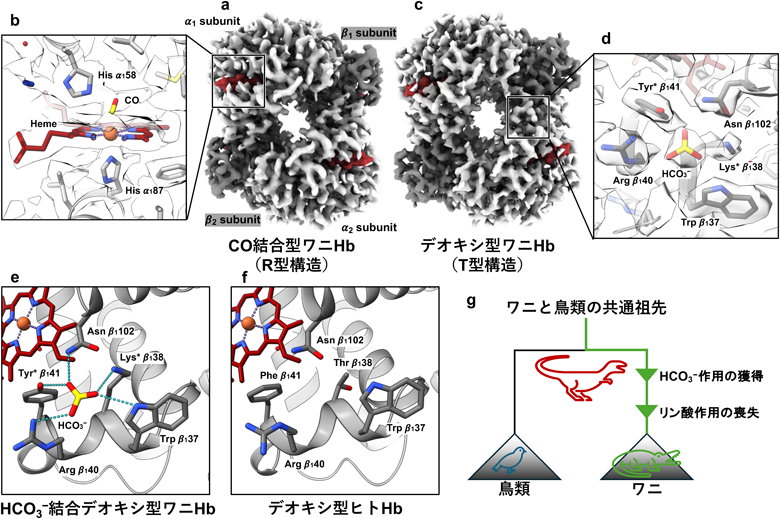
クライオ電子顕微鏡を用いた構造解析によるワニヘモグロビン特有のアロステリック制御の解明Cryo-EM structural analysis of crocodilian haemoglobin revealing its unique allosteric regulation
横浜市立大学大学院生命医科学研究科Graduate School of Medical Life Science, Yokohama City University ◇ 〒230–0045 神奈川県横浜市鶴見区末広町1–7–29 横浜市立大学鶴見キャンパス ◇ 1–7–29 Suehiro-cho, Tsurumi-ku, Yokohama, Kanagawa 230–0045, Japan
発行日:2025年8月25日Published: August 25, 2025