1)多細胞をロバストにする原理はいまだ不明である
多細胞生物が健康に生存するためには,発生過程では多種多様な細胞を秩序正しく配置し,成体では細胞を定期的に入れ替えつつ,その秩序と機能を維持する必要がある.ところが,こうした発生や新陳代謝は動的な生命現象であるため,秩序を脅かす撹乱によって頻繁に揺らぐ.たとえば,細胞の活発な分裂・移動・代謝は,複製エラーや情報伝達異常,細胞の配置ミス,細胞傷害誘発物質(活性酸素等)の産生など,細胞状態の揺らぎを頻繁に引き起こす.しかし,多細胞生物は,このような揺らぎを乗り越えて秩序正しい細胞配置とその機能維持を安定的になしとげる.具体例をあげると,組織の細胞配置決定を担う基幹システムであるモルフォゲン勾配は,細胞が活発に分裂・運動を行う組織(つまりモルフォゲン勾配が揺らぎやすい環境)であっても,それを乗り越えて正確な細胞配置を導く.また,ヒト胚では,着床前の段階で異数性細胞(染色体数が異常な細胞)が生じたとしても発生が進むと異数性細胞が次第に消失し,胎内で健康に発生する1).こうした,揺らぎを乗り越えて多細胞体の秩序を頑強(ロバスト)にする性質,「ロバストネス」は,あたり前のように生物学者に捉えられてきた.しかしながら,実は,多細胞体のロバストネスを実現する原理の理解はいまだに進んでいない.
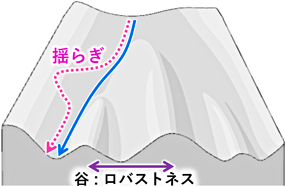
生物学的なロバストネスの概念は,前世紀中盤にC.H. Waddingtonにより提案された2, 3).彼が作成したランドスケープモデル(図1)は「細胞の運命が一方向に進み,多様化していくこと」を示す図としてよく使われるが,同時に“谷”でロバストネスを表現しており,「極端な揺らぎが生じた場合には運命が転換してしまうが,閾値を超えない範囲(地形の稜線まで)の揺らぎであれば運命が転換することなく(斜面を転がり戻るように)対処できる」という概念を示した.その後,分子生物学の進歩に伴い,発生を支えるロバストネスの分子実体として「遺伝的補償」や「熱ショックタンパク質によるタンパク質の構造・機能の補償」などが提案され,注目を集めた4–6).ところが,これらは「個々の細胞が単独で実現できるロバストネス」を説明するものであり,多細胞生命の本質ともいえる細胞間の連携が生体組織をロバストにする原理は依然として不明である.すなわち,性質や機能が異なる多様な細胞の集合体である生体組織をロバストにするには,それを構成する多様な細胞が連携して「組織全体の状態の感知と,それに応じた細胞集団の運命の時空間的制御」を実現する必要があるが,そのような細胞間連携の理解は進んでいない.また,従来の発生研究や疾患研究は,変異体や病態などの明確な表現型を頼りに進展してきたが,これに対し,ロバストネスは「生体内で偶発的に起こる揺らぎを解消するシステム」であるため,その実態を捕捉することさえきわめて困難であった.
2)細胞競合の生理的機能はほとんど不明である
近年,新たな細胞間連携システムとして,細胞競合が注目されている,細胞競合は,単独では生存できる異質な細胞が,正常細胞と共存した場合に集団から競合的に排除される現象である.その最初の発見は50年前であり,スペインのMorataらによって,リボソームタンパク質遺伝子に変異を持った細胞(単独では生存できるが機能が低下した細胞)をショウジョウバエ正常上皮組織に誘導すると組織から排除される現象として見いだされた7).しかし,その後はあまり注目されることなく長らく埋もれていたが,2009年に井垣達吏博士と藤田恭之博士がそれぞれショウジョウバエ上皮と哺乳類培養上皮において,前がん細胞が正常細胞に囲まれると排除されることを見いだし8, 9),細胞競合が「がん抑制」機構として働きうることと,哺乳類においても細胞競合が起こることを世界に先駆けて示した.これを契機に,細胞競合によるがん抑制機構の研究が爆発的に増え,Keystone Symposiaを含め5回の関連国際シンポジウムが開催される(2015, 2016, 2017, 2020, 2024年)など世界的に細胞競合研究が盛り上がり始めている.しかし,現状,日本を含めて世界で行われている細胞競合研究の大半は,人為的に誘導した異常細胞が誘起する競合現象を解析するにとどまっており,細胞競合の生理機能は不明なままであった.そのため,細胞競合は,「興味深い現象ではあるが,生理的な機能を持たない,人工的にしか見えない現象ではないか」という見方も少なくない状況であった.
そのような中で,筆者の研究チームは,これまで続けてきたモルフォゲンシグナル研究を深める過程で,「細胞競合が生理的に機能し,多細胞のロバストネスを支えること」を発見するに至った.続く節では,この発見について紹介する.
2. 生理的な細胞競合がモルフォゲン勾配のロバストネスを支える
1)モルフォゲン勾配のリアルタイム可視化に挑む
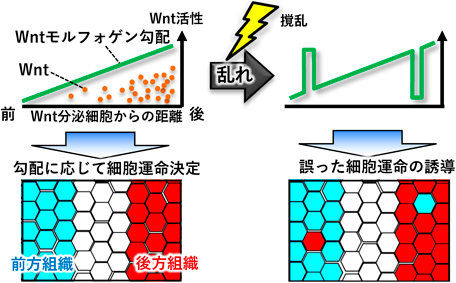
筆者は,モルフォゲンの一つであるWntに注目し,動物発生におけるモルフォゲンシグナル伝達機構と機能を長年研究してきた10–14).モルフォゲンは,生物の体あるいはそれを構成する組織にパターンを与える分子システムである.生物の体や組織が正常に機能するためには,特定の機能を備えた細胞を適切な位置に配置する必要がある.このような細胞配置(パターン)は,モルフォゲン勾配によって作り上げられる.モルフォゲンは発生源から濃度勾配を持って発せられ,その濃度に応じて異なる強さのシグナルを細胞に入力し,結果としてモルフォゲンシグナル強度の勾配(モルフォゲン勾配)が形成され,この勾配に沿って各細胞が自身の位置情報を把握し,その位置に適合する運命を選択する.一例をあげると,脊椎動物の初期胚の前後軸に沿ったパターンの形成は,Wntモルフォゲン勾配によって制御される(図2左).Wntモルフォゲン勾配が形成された初期胚では,後方に位置する細胞でWntシグナルが強く活性化され,その結果として,後方組織にふさわしい細胞(脊髄など)の前駆細胞に分化するための遺伝子発現変化が誘導される.一方で,前方に位置する細胞ではWntシグナルが活性化せず,結果,前方組織にふさわしい細胞(終脳など)の前駆細胞に分化するための遺伝子発現変化が誘導される.このようにして,初期胚の前後に沿った規則正しい細胞配置が作り上げられる(図2左下).このモルフォゲン勾配のコンセプトは,現在,広く受け入れられている.ところが,このコンセプトはモルフォゲンの発現パターンや,モルフォゲンおよびそのシグナルの機能改変実験をもとに提唱されたものであり,Wntモルフォゲン勾配の形成プロセスをしっかり可視化解析した研究はなく,「図に描いたようなモルフォゲン勾配が本当に形成されるのか」は実は不明であった.また,当時のモルフォゲン研究は,モルフォゲンの機能やモルフォゲンを受容する細胞内でのシグナル伝達機構の研究は精力的に行われてきたが,モルフォゲンを受容する細胞どうしのコミュニケーションについては研究されてこなかった.
そこで,筆者は,2006年に独立研究者になって以来,Wntモルフォゲンをモデルに,in vivoにおけるモルフォゲン勾配形成過程の高解像度での可視化に取り組んできた.まず,不安定化GFP(d2EGFP)を利用した高感度Wntシグナルレポーターを開発し,イメージングに適したモデル脊椎動物であるゼブラフィッシュにこれを組み込み,脊椎動物におけるWntモルフォゲンのシグナル動態の可視化に成功した15).このWntシグナル可視化ゼブラフィッシュは,多様な組織・臓器におけるWntシグナル動態を高感度に可視化できるため,現在,世界中の発生・再生研究者に使用されており,ナショナルバイオリソースプロジェクトゼブラフィッシュ(日本のゼブラフィッシュ系統を収集して世界の研究者へ配布する組織)の中でも「歴代ベストセラー商品」の一つとなっている.しかし,蛍光タンパク質を利用したこのレポーターには,ダイナミックレンジの不足,リアルタイム性の不足,自家蛍光の存在,といった欠点があった.そこで続いて,これらの弱点を解消できる新たなレポーターの作製に挑戦してきた.その結果,ブラジル産ヒカリコメツキムシ由来のルシフェラーゼElucに分解配列を付与し,Wntシグナルの入力直後から入力強度に比例した強さで発光し,シグナルが止まると15分以内に発光停止する,発光型リアルタイムレポーターの作製に成功した16).発光のイメージングは蛍光と違って技術が発展途上で解析に苦戦したが,オリンパス社(現・EVIDENT社)のご助力を得て安定的に可視化することに成功し,上述した初期胚前後軸に沿ったWntモルフォゲン勾配を観察することができた16).
2)モルフォゲン勾配は頻繁に乱れる
このように,これまでにない高い感度とリアルタイム性を持つレポーターを独自に構築することができ,満を持してゼブラフィッシュ胚のモルフォゲン勾配の形成プロセスを詳細に解析した.その結果,奇妙なことに気づいた.それは,野生型胚であるにもかかわらず,Wntシグナル強度が弱くあるべき前方組織でシグナル強度が異常に高い細胞が生じたり,逆にシグナル強度が強くあるべき後方組織でシグナル強度が異常に低い細胞が生じたりすることで,頻繁にWntモルフォゲン勾配が乱れてしまう,という現象である(図2右).このような異常な細胞(ノイズ細胞)が残存してしまうと誤った細胞運命が誘導されて発生異常が生じかねないが,興味深いことに,このようなノイズ細胞が生じた胚であっても最終的には正常に発生した16).この事実は,ゼブラフィッシュ胚がこのモルフォゲン勾配のノイズを何らかの方法で解消している可能性を示している.
3)モルフォゲン勾配を乱すノイズ細胞は細胞死により除去される
では,どうやって,モルフォゲン勾配のノイズが解消されるのだろうか? 我々は,慎重なイメージング解析を行い,その結果,ノイズ細胞が細胞死(アポトーシス)を起こして組織から除去され,結果としてモルフォゲン勾配の乱れが修復されることがわかってきた16).さらに興味深いことに,ノイズ細胞をゼブラフィッシュ胚に人為的に導入した場合においてもノイズ細胞がアポトーシスを起こした.具体的には,Wntシグナルの構成因子(図3)を強制活性化あるいは不活性化した細胞をゼブラフィッシュ胚へ導入した.まず,Wntシグナル構成因子βカテニンを恒常的に活性化させた細胞(Wntシグナル異常活性化細胞)を導入するとWntシグナル活性がもともと低い胚前方領域ではアポトーシスを起こしたが,一方で,シグナル活性がもともと高い後方領域にそれらを導入してもアポトーシスが起きなかった.また逆に,シグナル異常低下細胞(Wntシグナルの負の制御因子であるAxinあるいはGSK3βを恒常的に活性化させた細胞)を導入した場合は,シグナル活性がもともと高い後方領域でアポトーシスを起こした.すなわち,場に不適合なモルフォゲンシグナル活性を持つ細胞が選択的に排除されることで,モルフォゲン勾配の乱れが修復されることがわかった16).
4)カドヘリンがモルフォゲンノイズ細胞の排除に関わる
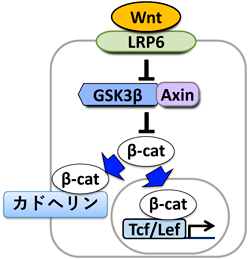
上述の発見により,「モルフォゲン受容細胞が互いにモルフォゲンシグナル活性を監視しあっており,場に不適合なシグナル活性を持つ細胞が出現すると,細胞競合によってそれを積極的に排除する可能性」がみえてきた.しかしながら,この時点では,モルフォゲン受容細胞間のコミュニケーションは知られておらず,この現象を細胞競合であると結論づけることはできなかった.そこで我々は,「隣接し合うモルフォゲン受容細胞が互いのシグナル活性を監視しあう機構」を探っていった.まず,Wntシグナルの情報伝達のどのステップの異常がノイズ細胞排除を起動するのかを調べた.正常にWntシグナルを伝達する細胞においては,モルフォゲン分子Wntが受容体群(LRP6など)を介して細胞内にシグナルを入力し,GSK3βやAxinからなる分解複合体を不活性化することで細胞質中のβカテニンタンパク質の安定化を導く.そして,安定化したβカテニンが核内に移行して転写因子Tcf/Lefを活性化し,標的遺伝子の発現を導く(図3).我々は当初,核内での転写活性の異常がノイズ細胞排除を起動すると予想していた.ところが,予想外なことに,Tcf/Lefを強制活性化あるいは不活性化した細胞を導入しても排除が起きず,一方で,LRP6やβカテニン,GSK3βなどを強制活性化あるいは不活性化した細胞は排除された.また,核外移行シグナルを付与したβカテニンを過剰発現させた細胞を導入すると排除されたが,核内移行シグナルを付与したβカテニンを過剰発現させた細胞を導入しても排除されなかった16).これらの事実は,核外のβカテニンの量の異常がノイズ細胞排除を導く可能性を示している.
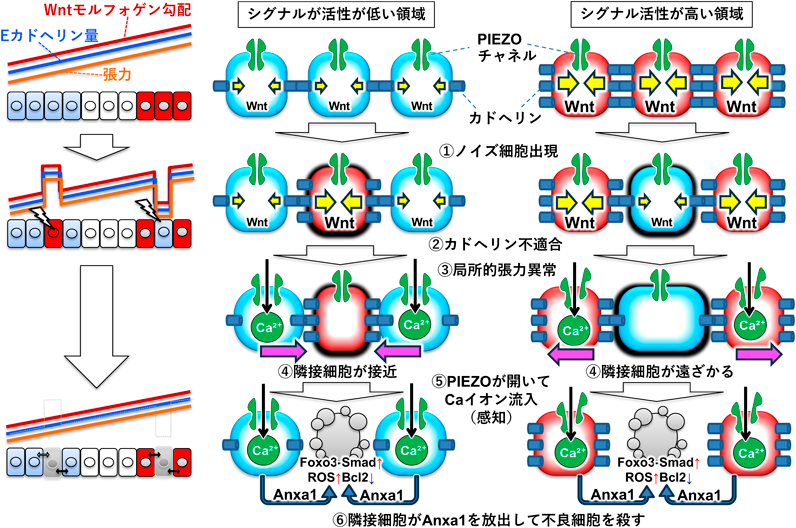
ここまでの解析結果を受け,我々は次に,βカテニンの“もう一つの機能”に注目した.βカテニンは,Wntシグナルの標的遺伝子発現を制御するのみならず,細胞膜で細胞接着タンパク質カドヘリンと相互作用する(図3).カドヘリンは,細胞外ドメインを介してそれどうしで結合する能力を持っており,カドヘリンを膜上に発現する細胞は,カドヘリンを介して接着することができる.我々は,ゼブラフィッシュ胚においてWntシグナルが核内のβカテニンのみならず細胞膜のβカテニンをも安定化し,Wntシグナルによって安定化された細胞膜βカテニンがEカドヘリンの膜局在を促進すること,そしてその結果として,Wntシグナル活性の高い後方組織の細胞ではその細胞膜上に多数のEカドヘリン分子がリクルートされ,一方,Wntシグナル活性の低い前方組織ではEカドヘリンのリクルートが起こらず,膜上のEカドヘリン分子の数が少ない状態になることを見いだした(図4左上).また,場に適さないWntシグナル活性を持ったノイズ細胞では,シグナル活性に応じてカドヘリン量が変化するため,結果としてノイズ細胞と隣接正常細胞の間でカドヘリン量の不適合が生じた(図4①②).このカドヘリン量の不適合がノイズ細胞の排除を駆動することがわかってきた.実際,胚全体でカドヘリンの量を強制的に増加あるいは減少させると,Wntシグナルが不適合なノイズ細胞を誘導しても排除が起きなくなり,また,カドヘリン量が場に不適合な細胞を正常胚に導入すると排除された16).このように,ゼブラフィッシュ胚の細胞が,この「Wntシグナル–カドヘリン変換」を利用して隣接細胞のWntシグナル活性を測ることで,ノイズ細胞を検出し,その排除を起動することがわかってきた.また,これらの結果から,この「自然発生した異質な細胞が正常細胞との相互作用を経て排除されることでモルフォゲン勾配が修復される現象」が生理的な細胞競合である可能性がみえてきた.
5)局所的張力変動がモルフォゲンノイズ細胞の排除を起動する
続いて我々は,カドヘリン量の不適合がどのようにしてノイズ細胞の排除を起動するのかを調べた.カドヘリンは,細胞外では隣接細胞のカドヘリンと結合することで細胞接着を形成するが(図4上段),細胞内ではアクトミオシン(アクチンミオシン複合体)と結合することで引っぱる力(張力)を発生させる.このため,カドヘリンを介して結合する細胞と細胞の間には互いを引っぱり合う力,細胞間張力が生じる(図4上段,細胞内の矢印).我々は,この事実をもとに「Wntシグナルの強さがカドヘリン量に変換され,最終的に張力に変換される」という仮説を立て,ゼブラフィッシュ胚で検証を行った.張力マーカー(RhoA活性やミオシン軽鎖リン酸化,Fアクチン集積など)の可視化や,組織をレーザーで切断して細胞間張力を測定する解析を行った結果,この仮説が正しく,Wntシグナル活性が高い領域では強い張力が発生し,活性が低い領域では張力が弱いことがわかってきた(図4上段).
さらなるイメージング解析の結果,このシグナルの張力への変換がノイズ細胞の感知に関わることも明らかになった(図4).具体的には,①場に適さないWntシグナル活性を持ったノイズ細胞が生じると,②その細胞でカドヘリン量が変化した結果として,③張力が異常変化し,局所的な張力異常が生じること(シグナルが高すぎると張力が異常に増え,低すぎると張力が異常に低下すること),ノイズ細胞で張力異常が生じた結果として隣接細胞との細胞間張力のバランスが崩れ,④隣接細胞が不良細胞に引き寄せられ(あるいは遠ざかり),この急激な隣接細胞の動きがストレスとなって,⑤細胞膜の物理的ストレスによって開閉するカルシウムイオンチャネルPIEZO1が開いてカルシウムイオンが隣接細胞内に流入することがわかった.また,PIEZO1をノックアウトあるいは薬剤で阻害すると,ノイズ細胞の細胞死が抑制された.したがって,ノイズ細胞の張力変化の結果として起こる隣接細胞におけるPIEZO1の活性化によって,ノイズ細胞排除が起こると考えられる16).
6)隣接細胞からのAnxa1がノイズ細胞の細胞死を誘導する
では,PIEZO1の活性化はどのようにして,ノイズ細胞除去を引き起こすのだろうか? これまでの細胞競合研究では,ショウジョウバエ遺伝学などによってメカニズム解析が進んできたが,我々が用いるゼブラフィッシュの系で同様な遺伝学スクリーニングを行うと膨大な労力と時間がかかってしまう.ゼブラフィッシュをモデルとした生理的細胞競合の解析という未踏の課題にどう挑むか,非常に悩んだが,九州大学生体防御医学研究所教授(現・所長)の大川恭行博士のご助言とサポートを得て,独自のトランスクリプトーム解析系を立ち上げた.具体的には,蛍光ラベルしたノイズ細胞をゼブラフィッシュ胚に導入し,FACS(fluorescence-activated cell sorting)でノイズ細胞とそれ以外の細胞(ノイズ細胞の隣接細胞を含む正常細胞群)を分取し,トランスクリプトーム解析を行った.そして,まず,単純なWntシグナルの増減では発現変化する遺伝子を取り除き,続いて,「ノイズ細胞特異的に発現が増減する遺伝子群」や「ノイズ細胞の出現依存的に正常細胞で発現が増減する遺伝子群」を制御因子候補として絞り込んだ.そしてこれらの候補群の解析の結果,PIEZO1の下流で分泌タンパク質アネキシンA1(Anxa1)の発現が誘導され,Anxa1がノイズ細胞における転写因子Foxo3と転写因子Smad(TGF-βタイプのSmadファミリーであるSmad2/3/4)の活性化を介して活性酸素(ROS)の産生を誘導すること,そして産生されたROSが細胞死抑制因子Bcl2の分解を誘導し,ノイズ細胞のアポトーシスを誘導することが明らかになった(図4)16–18).また,ノイズ細胞の感知・排除における必須のプロセスであるPiezo1活性化やAnxa1発現誘導,ROS産生,Bcl2減少を抑制すると,いずれのケースにおいても,自然発生したノイズ細胞が蓄積し,Wntモルフォゲン勾配が乱れ,また,前方領域に後方領域に作られるべき脊髄の前駆細胞が生じたり,後方領域に前方領域に作られるべき終脳の前駆細胞が生じたりするなど,位置情報があべこべになった16, 17).このように,モルフォゲン受容細胞の細胞競合によってモルフォゲン勾配とそれによって誘導される組織パターンのロバストネスが支えられることが明らかになった.また,この一連の研究により,細胞競合が生理的に働く,発生ロバストネスを支えるシステムであることが示すことができた.
3. 生理的細胞競合が多様な組織のロバストネスを支える
1)生理的細胞競合が多様な異常細胞を排除する
我々の研究と並行し,2019年に大阪大学生命機能研究科の佐々木洋博士らが,マウス初期胚において「多能性が低い細胞が自然発生するものの細胞競合によって排除されること」を報告した19).さらに,2021年にはRodriguezらによって,マウス初期胚が生み出す細胞のうち35%もの細胞が異常細胞として細胞競合によって除去されており,それらの多くにおいてミトコンドリア活性異常が生じていることが報告された20).このように,生理的細胞競合が「胚に生じた多様な異常細胞を排除して発生ロバストネスを支えるシステム」であることを示すエビデンスが蓄積されつつある.
2)生理的細胞競合が脊髄や筋肉のロバストな形成を支える
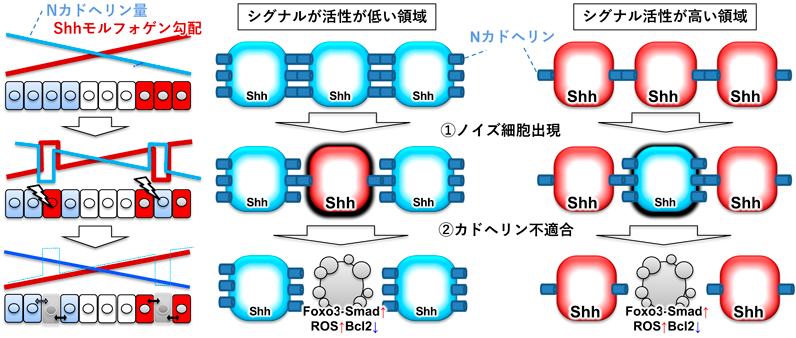
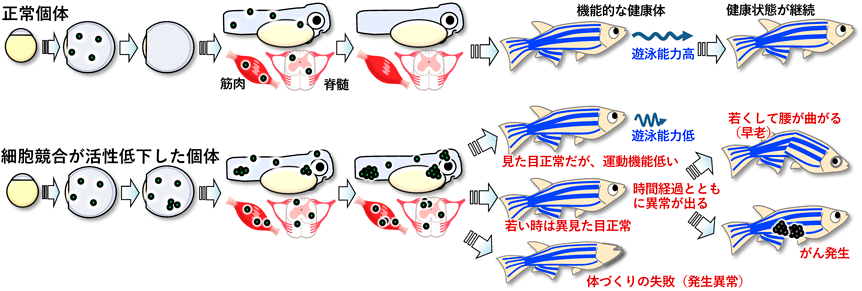
上述したように,生理的細胞競合が胚発生のロバストネスを支えることがわかってきているが,一方で,臓器の構築などの発生後期のプロセスのロバストネスにも細胞競合が関わるのかはまったく不明であった.そこで,筆者らは,Shhモルフォゲンによってパターン形成される筋肉と脊髄に注目した.モルフォゲン分子Shhは,形成過程の脊髄と筋肉において,腹側領域から分泌されて背腹軸に沿ってShhシグナルの活性勾配(Shhモルフォゲン勾配)を作り出す.そして,そのシグナル入力強度に応じて,脊髄原基(神経管)においては異なるタイプの運動神経細胞と介在神経細胞,筋肉原基においては速筋細胞と遅筋細胞の形成を誘導する.つまり,適切な位置で適切な強度でShhシグナルが活動することにより機能的な脊髄と筋肉が形成される.まず,我々は,ゼブラフィッシュにおいてShhシグナルの活動を可視化し,その結果,筋肉と脊髄の形成過程において本来Shhシグナルの活性が低く抑えられるべき領域でShhシグナルが異常に活性化した細胞が生じたり,逆にShhシグナルが強く活性化される領域でShhシグナルが活性化しない細胞が生じたりしてしまうことを確認した(図5中段).また,メカニズム解析の結果,Wntモルフォゲン勾配のノイズ解消と類似した細胞競合機構によってShhモルフォゲン勾配のノイズが解消されることがわかってきた.まず,Shhシグナルがその強度に応じて細胞膜のNカドヘリン量を減らすため(WntシグナルはEカドヘリン量を増やす),結果として,Shhシグナル活性が高い領域では細胞膜上のNカドヘリン量が少なく,活性が低い領域ではNカドヘリン量が多くなる(図5上段).その結果,突発的に生じたShhシグナルノイズ細胞ではNカドヘリン量が変化し(図5中段),この変化を隣接正常細胞が感知し,ノイズ細胞におけるFoxo3とSmadの活性化,ROS産生,Bcl2分解を誘導して,ノイズ細胞を細胞死させる(図5下段).また,Foxo3活性化やROS産生,Bcl2減少を抑制した(細胞競合を阻害した)ゼブラフィッシュでは,胚や筋肉・脊髄において自然発生したWntシグナル異常細胞やShhシグナル異常細胞が蓄積して運動神経細胞や介在神経細胞,速筋・遅筋細胞が間違った場所に配置され,さらに稚魚に成長後,一部の個体は発生異常(目が欠損したり尻尾が曲がったり体長が短かったり)を示し,また別の個体は,見た目が正常にもかかわらず遊泳能力が低かったり,時間の経過とともに腰が曲がったり腫瘍ができたりなど早老の異常を示した(図6)18).異なるタイミングで異なる部位に発生した異常細胞が細胞競合破綻により蓄積した結果として,このような多様な表現型が生じたと推察される.このように,生理的な細胞競合が速筋・遅筋細胞や脊髄内の神経細胞の正確な配置,運動機能の発達に必須であること,すなわち,生理的細胞競合が初期胚発生のみならず器官構築のロバストネスも支え,健康的な個体の形成に必須であることが明らかになった.
3)ゼブラフィッシュ生体内で多様な細胞競合が働く
上述のように,これまでの細胞競合研究はショウジョウバエ上皮と哺乳類培養上皮をモデルに発展し,リボソーム遺伝子変異細胞や前がん細胞,Myc低発現細胞などが細胞競合によって排除されることを明らかにしてきた.しかし,これらの系では,脊椎動物生体内における細胞競合を理解するのは困難であった.そのような中で筆者らは,上述のように,ゼブラフィッシュをモデルとして脊椎動物生体内における細胞競合解析系を立ち上げ,場に不適合なWnt/Shhシグナル活性を持つ細胞を排除する新たな細胞競合現象とその分子機序の解明に成功した16–18).この経緯から我々は,既知の細胞競合現象についても同様に,ゼブラフィッシュをモデルに脊椎動物生体における制御機構を解析できるのではないかと考えた.そして,これまでに,ゼブラフィッシュにおいても前がん細胞(Ras変異細胞やSrc活性化細胞)やMyc低発現細胞,リボソーム遺伝子変異細胞が細胞競合によって排除されることを確認しており,この系を駆使して,Ras変異細胞が隣接正常細胞に細胞老化を誘導されるなど,従来の培養細胞解析ではみえなかったメカニズムを明らかにしつつある16–18, 21).
4. 細胞競合マーカーが生理的細胞競合研究の未来を拓く
1)細胞競合マーカー研究の現状
上述のように,生理的細胞競合が発生中の脊椎動物個体内に生じたエラーを解消する一種の生体防御システムであることが明らかになりつつあるが,初期胚や脊髄・筋肉原基などごく限られた組織でしか生理的細胞競合の活動は捕捉されておらず,他の器官構築過程や成体組織の再生・老化プロセスにおいても生理的細胞競合が機能しているのかはよくわかっていない.発生・再生する個体全身の中で突発的に生じる病的細胞を捉えることは困難であり,それゆえ,生理的細胞競合の機能を探るのは困難な状況である.したがって,今後,生体内における細胞競合の機能を理解していくためには,脊椎動物体内で起こる「生理的細胞競合を捕捉するためのマーカー分子」が有効なツールになるのは間違いない.これまで,細胞競合マーカーの探索はショウジョウバエにおいて進められおり,現在までにFlower LoseとAzotが細胞競合マーカーとして同定されている.そして,これらを利用した解析により,ショウジョウバエにおける生理的細胞競合が寿命延伸に必須であることなどが示されている22, 23).しかしながら,ゼブラフィッシュやマウスの胚の生理的細胞競合プロセスではFlower LoseやAzotの脊椎動物ホモログの発現変化は検出されず18),脊椎動物の生理的細胞競合マーカー分子は不明なままであった.その上,脊椎動物において細胞競合は多様な異常性(Wntシグナル異常,Shhシグナル異常,Myc低発現,多能性低下,ミトコンドリア機能異常,リボソーム遺伝子異常,発がんシグナル活性化など)によって起動されるため,それを感知・排除するメカニズムも多様であることが予測され,すべての細胞競合を捕捉できるような分子を見つけることは不可能に近い,とも考えられてきた.
2)細胞競合マーカー同定への挑戦
上記3節3項で言及したように,我々はゼブラフィッシュにおいて多様な細胞競合現象を解析することを可能にしている.近年,細胞競合研究者は増えているが,我々のように多様な細胞競合を解析できる研究グループはまれであり,この環境を活かすことで,細胞競合マーカー同定という未踏の課題に挑戦できると考えた.まず我々は,Wntシグナルノイズ細胞とShhシグナルノイズ細胞の排除メカニズムの共通制御因子を探っていった.その結果,WntシグナルあるいはShhシグナルが場に適さず強すぎる細胞または弱すぎる細胞(細胞競合の敗者となる四つの異なる条件)で共通して発現上昇する遺伝子として,foxo3, sesn3, lnx, tcimaという四つの遺伝子を見いだした.さらに,このうちfoxo3が,細胞競合の敗者となる前がん細胞やMyc低発現細胞,リボソーム異常細胞でも発現誘導され,加えて,マウス胚において敗者細胞となる多能性低下細胞でも発現誘導されることがわかってきた18).また,上述したように,Foxo3が敗者細胞の排除に必須であることも判明した.このように,複数の細胞競合で共通して活性化し,かつ,種を超えて働く,細胞競合の敗者細胞マーカーとしてfoxo3を同定することができた.
3)foxo3をマーカーとした細胞競合の可視化
細胞競合で排除されるような異常細胞は不規則に生じるため,結果として,それが生体内のどこにいつ生じるのか,そしてそれが駆動する細胞競合がいつどこで起こるのかは,これまでは捕捉困難であった.しかし,細胞競合の共通マーカーとしてfoxo3を発見したことにより,生体内における細胞競合の動態を一挙に把握できる可能性が出てきた.そこで我々は,foxo3の発現を高感度に検出できるレポーターゼブラフィッシュを作製し,発生プロセスにおけるfoxo3の発現動態を観察した.その結果,初期胚発生や筋肉・脊髄の形成過程においてランダムにfoxo3発現細胞が出現し,アポトーシスを起こして消えていく様子を捉えることができた18).現在,このレポーターを駆使して未知の生理的細胞競合の機能と制御の解析を進めている.
4)細胞競合マーカーの残された課題
さて,foxo3を細胞競合マーカーとして同定することができたが,一方で,foxo3は細胞競合以外の生命現象においても活性化されることも知られており,そのため,foxo3は細胞競合の「共通マーカー」であるとはいえても「特異的マーカー」とまではいえない.そこで我々は現在,この課題を解決するために二つのアプローチを進めている.一つは,細胞競合の勝者細胞マーカーの同定,もう一つは,細胞競合の敗者細胞あるいは勝者細胞に特異的に発現するマーカーの同定である.少なくとも,現時点までに,空間トランスクリプトーム解析によって複数の勝者細胞マーカー候補の同定に成功しており(未発表,論文作成中),敗者細胞の共通マーカーであるFoxo3と組み合わせることで,生理的細胞競合のより精度の高い可視化を可能にしつつある.
1)未知の生理機能の発見
上述のような高精度な細胞競合の可視化によって,今後新たな細胞競合の生理機能が明らかになるはずである.また,脊椎動物の生理的細胞競合の制御因子がわかってきたことにより,これらの機能改変実験によっても細胞競合がどのような細胞を排除しうるのかもわかってくるはずである.関連して興味深いのは,細胞競合制御因子Foxo3のヒトにおける遺伝子多型(遺伝子配列の違い)がヒトの個人差に関連する点である.具体的には,Foxo3の遺伝子多型が,身長差24),知能の差25),寿命の長さ26)など多様な個体差と関連することがわかっている.一つの可能性として,細胞競合の活性化度合い(Foxo3の活性の強さ)によって,特定の異常細胞を除去できなかった結果としてこうした個体差が生じたのかもしれない.もしかしたら,動物の一生涯における細胞競合の機能を調べていくことで,こうした個体差が生じるメカニズムの理解にもつなげていけるかもしれないと考えている.
また,細胞競合とモザイク疾患との関連についても検討していくべきである.出生後に生じた変異細胞に起因する疾患としては,「がん」がよく知られているが,近年のシングルセル解析技術の発展によって,アルツハイマー病や,てんかん,自閉症などの多様な疾患が,胚発生期に生じた変異細胞が生存・増殖して体細胞モザイクを形成することで引き起こされるケースがあることが明らかになっている27–30).したがって,これらの変異細胞が細胞競合によって排除されずに生存・増殖した可能性が考えられる.しかし,細胞競合の破綻の結果として変異細胞が生存したのか,あるいは変異細胞が細胞競合に耐性を持っているのかは不明である.現在までのゼブラフィッシュをモデルとした解析によって,神経変性疾患関連遺伝子に変異が生じた細胞が胚発生期に自然発生することと,それらが生理的な細胞競合によって除去されること,そして,特定の環境ストレスの存在下では細胞競合が破綻して神経変性疾患関連変異細胞が蓄積することがわかりつつある(投稿中).今後,個体の発生・成長過程で生じた変異細胞に起因する多様な疾患の発症と細胞競合,そしてそれらと環境因子の関連を調べることで,疾患発症の新しい原理が理解できるようになると期待している.
いま世界で注目されている「オートファジー」や「細胞老化」も,かつては生物学的意義が不明であったが,マーカーが同定されたことでその実態に取り組むことが可能になり,急速に分野が拡大した.細胞競合もこれらと同様にこれまでは生理的な意味がない現象かもしれないと軽視され,いまだに教科書にもほとんど記載されない状態であるが,我々による「発生ロバストネスを支えるシステムとしての脊椎動物における生理機能の報告(2019年)」と「メカニズムの詳細解明とマーカー同定(2024年)」が新たなブレークスルーとなり,ここから生理的細胞競合の研究が一気に進む(競争も激化するだろうが分野は間違いなく発展する)と期待している.
2)細胞競合を超えて,多細胞のロバストネス機構の理解へ
さて,私はモルフォゲンの研究を行う過程で“思いがけず”細胞競合の研究分野に飛び込むことになったが,とはいえ,やはり私の興味の根元にはモルフォゲンや多細胞のロバストネスがある.多細胞のロバストネスの原理を理解することを目標に置いたとき,細胞競合は重要な切り口となるが,一方で細胞競合だけではすべてを説明することは難しいだろう.細胞競合は,多細胞体が生み出した細胞を殺して減らすという「コスト的にむだが多そうにみえるシステム」であり,生じた異常細胞の異常の種類や程度によっては別の細胞間コミュニケーション機構が働いてしかるべきであろう.実際,最近になって,細胞競合同様に,場にフィットしない誤配置細胞とフィットしている近接細胞が連携しながら状態を最適化する現象「セルソーティング」が注目されている.具体的には,ゼブラフィッシュの脊髄前駆細胞の配置決定において前駆細胞の誤配置が生じると,近接細胞が誤配置細胞とのカドヘリンを利用した相互作用を経てその配置異常を感知し,誤配置細胞を生かしたまま適合部位に移動させることが2020年に報告された31).このセルソーティングは古くから議論されてきたが,この研究で初めて,発生ロバストネスを支える機構であることが示された.重要なことに,我々は,細胞競合とセルソーティングがクロストークしており,脊髄原基において一方を強制的に抑制すると他方がより活性化することを見いだしている18).我々は,これらの事実から,近接細胞間の積極的な連携が多細胞体のロバストネスを支える重要な原理であり,一方で,ロジックは共通しつつも異常(撹乱)の解消の方法が異なる複数のロバストネス機構が存在すること,そして,異常の程度や種類に応じて適切なロバストネス機構を選択することで多細胞体のロバストネスが実現するのではないか?と考えている.今後は,細胞競合研究を継続するとともに,こうした「未知のロバストネス機構」の探索と,複数のロバストネス機構の「選択・起動原理」を理解することで,多細胞ロバストネスを実現する細胞間コミュニケーション原理を解明していきたい.