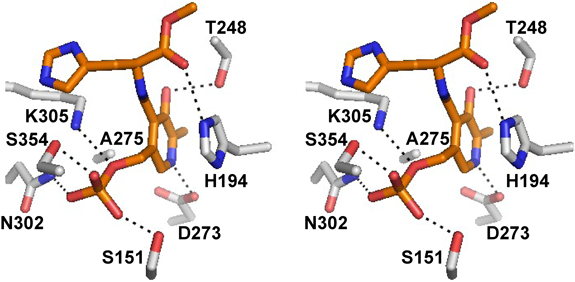
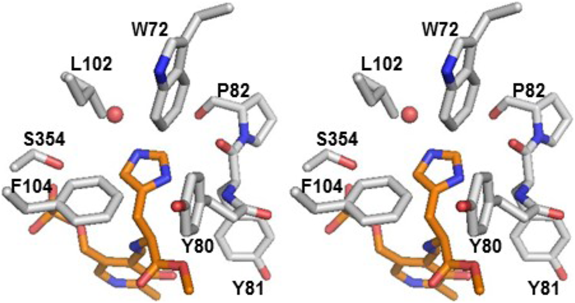
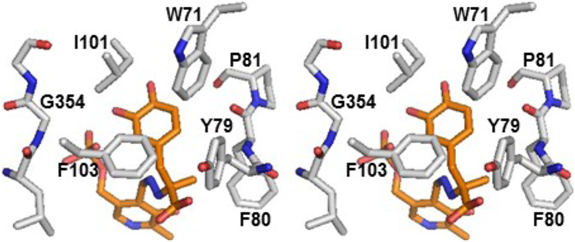
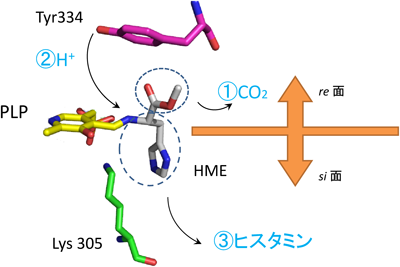
ヒスタミンとドーパミン生成酵素の変遷Relationship between histamine and dopamine synthesizing enzymes
1 岡山県立大学保健福祉学部Faculty of Health and Welfare Science, Okayama Prefectural University ◇ 〒719-1197 岡山県総社市窪木111番地111 Kuboki, Soja-shi, Okayama 719-1197, Japan
2 香川大学教育学部Faculty of Education, Kagawa University ◇ 〒760-8522 香川県高松市幸町1番1号1-1 Saiwai-cho, Takamatsu-shi, Kagawa 760-8522, Japan
3 龍谷大学農学部Faculty of Agriculture, Ryukoku University ◇ 〒612-8577 京都府京都市伏見区深草塚本町67番地67 Fukakusa Tsukamoto-cho, Fushimi-ku, Kyoto-shi, Kyoto 612-8577, Japan
発行日:2015年6月25日Published: June 25, 2015