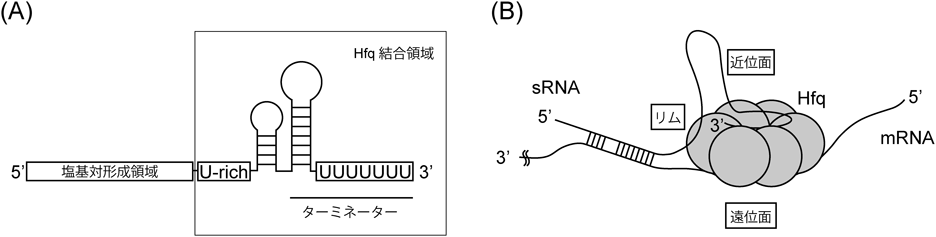
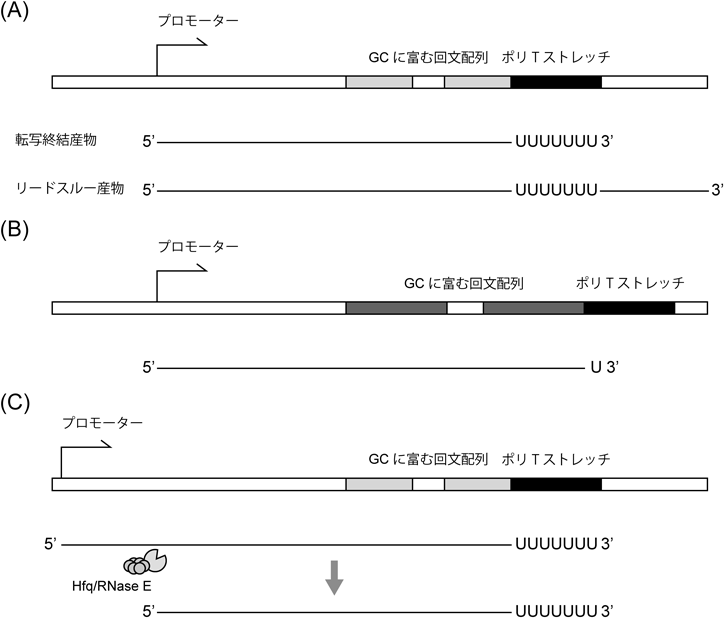
細菌小分子RNAの機能構造と生合成Functional structure and biogenesis of bacterial small RNA
鈴鹿医療科学大学薬学部薬学科Faculty of Pharmaceutical Sciences, Suzuka University of Medical Sciences ◇ 〒513–8670 三重県鈴鹿市南玉垣町3500–3 ◇ 3500–3 Minamitamagaki, Suzuka, Mie 513–8670
発行日:2017年8月25日Published: August 25, 2017