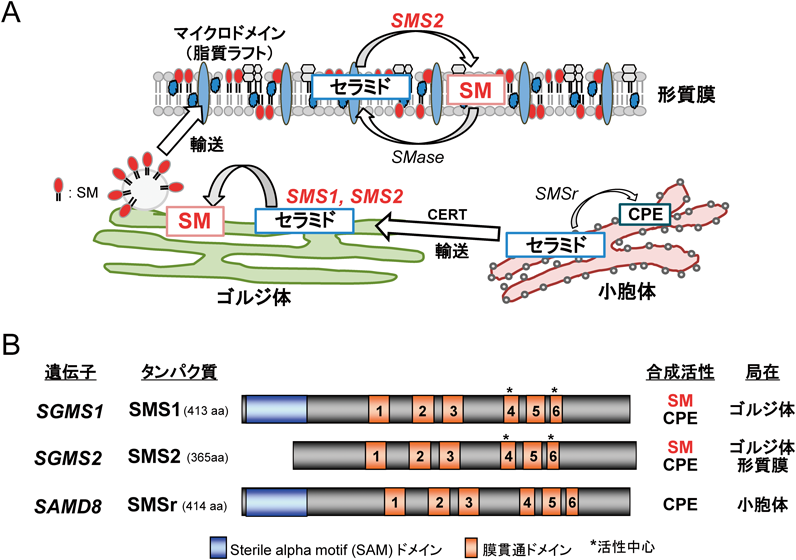
ウイルス感染におけるスフィンゴミエリン生合成の役割Role of sphingomyelin and its synthesis on viral lifecycle
金沢医科大学総合医学研究所生命科学研究領域Department of Life Science, Medical Research Institute, Kanazawa Medical University ◇ 〒920–0293 石川県河北郡内灘町大学1丁目1番地 ◇ 1–1 Daigaku, Uchinada, Ishikawa 920–0293, Japan
発行日:2018年4月25日Published: April 25, 2018