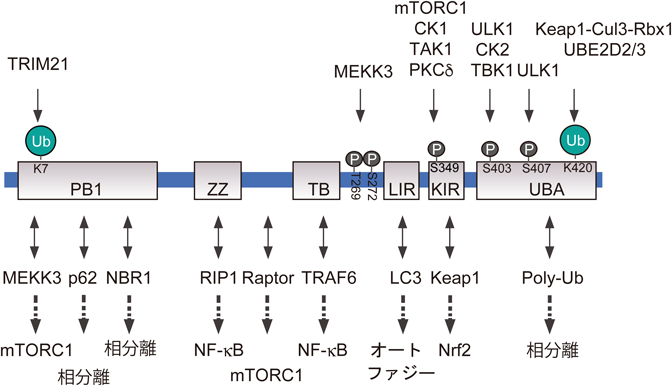
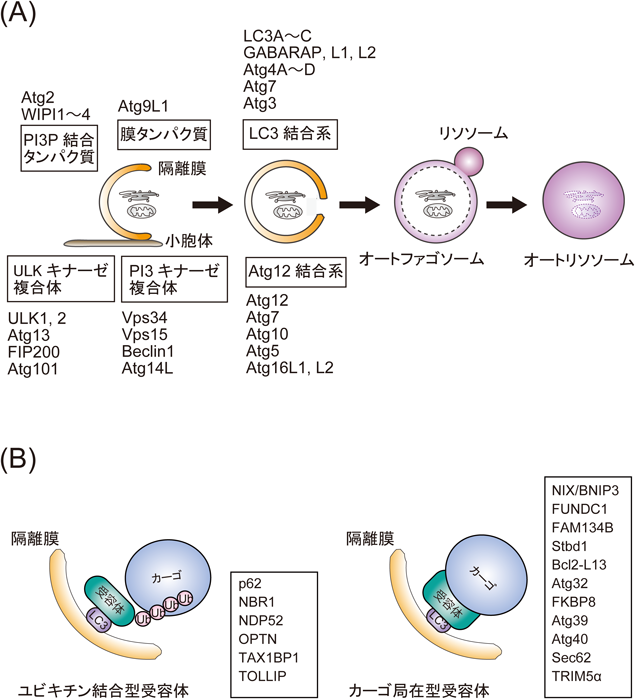
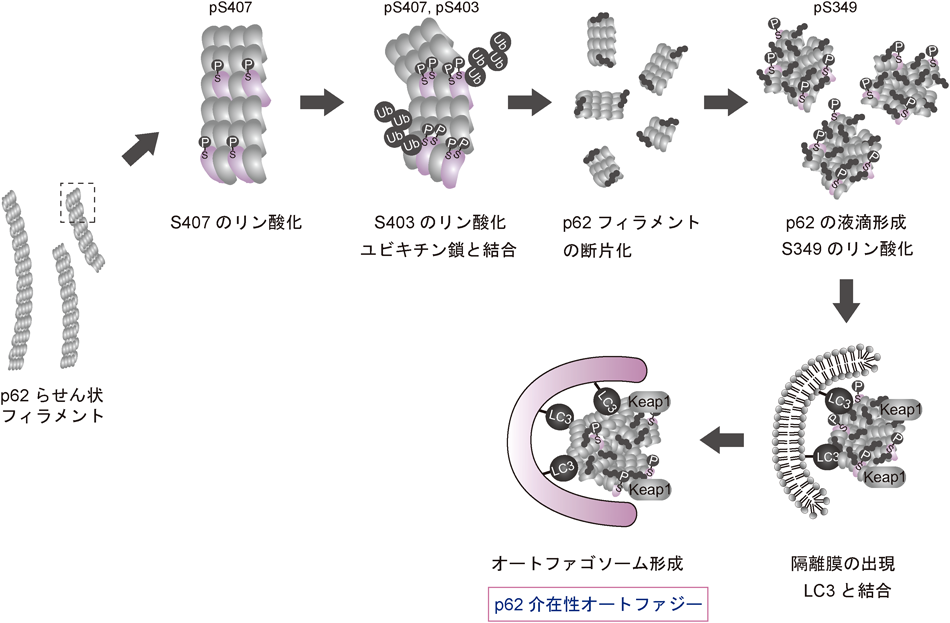
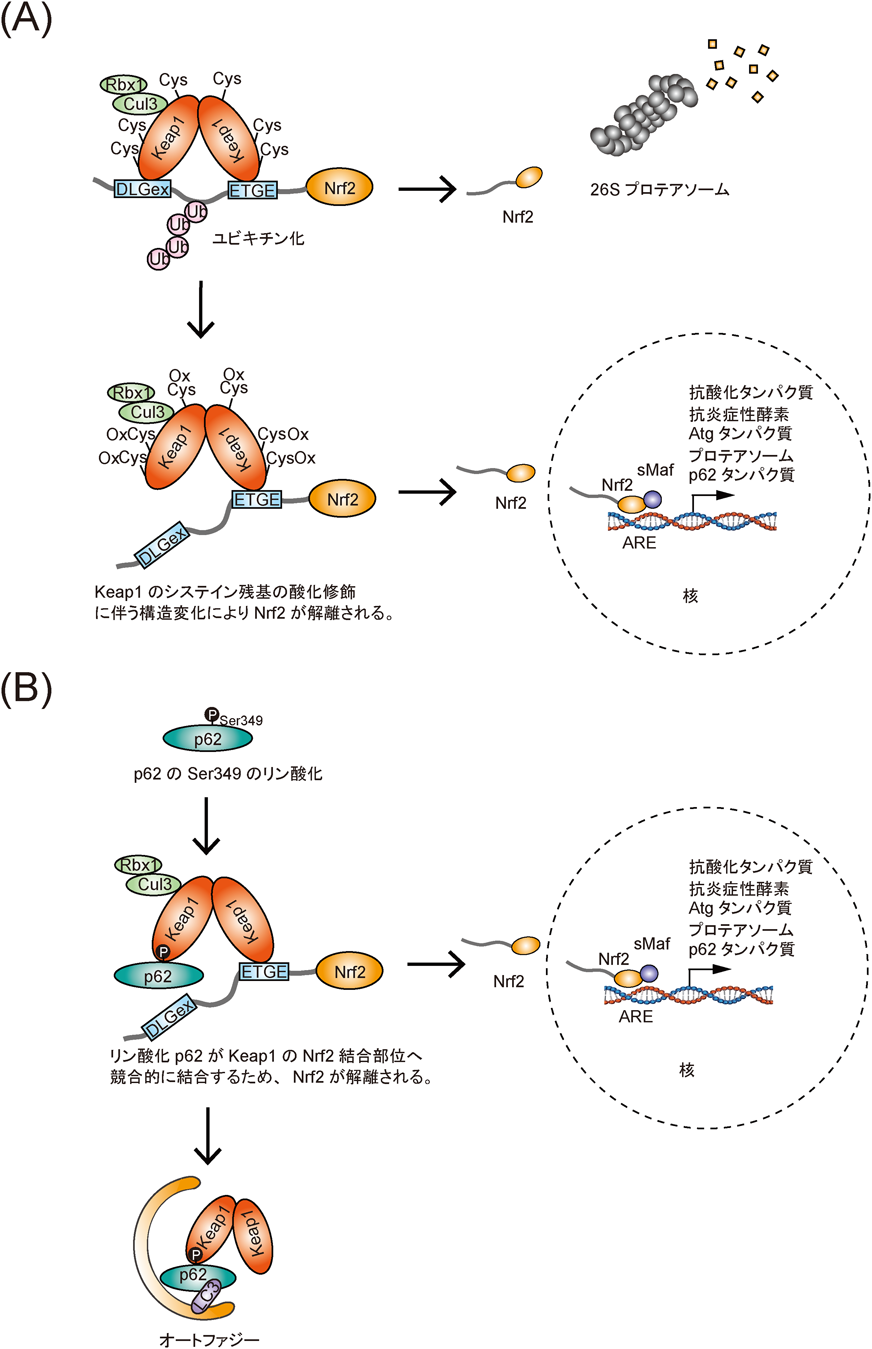
p62/SQSTM1 :オートファジーにおけるその役割Roles of p62/SQSTM1 in selective autophagy
順天堂大学医学部生理学第二講座Department of Physiology, Juntendo University Graduate School of Medicine ◇ 〒113–8421 東京都文京区本郷2–1–1 ◇ Hongo 2–1–1, Bunkyo-ku, Tokyo 113–8421, Japan
発行日:2019年6月25日Published: June 25, 2019