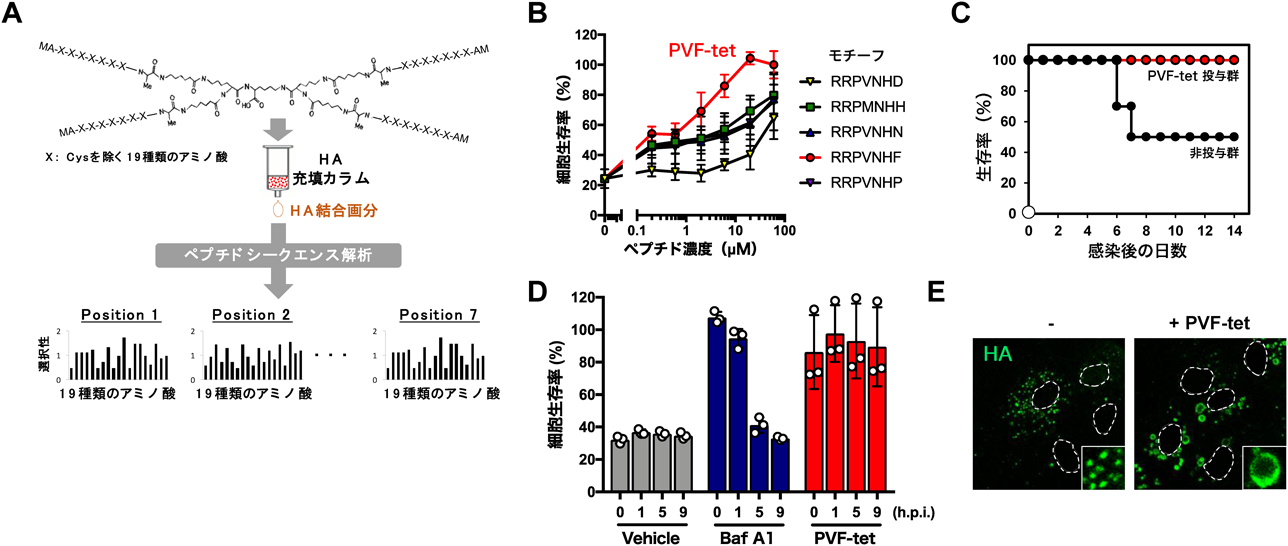
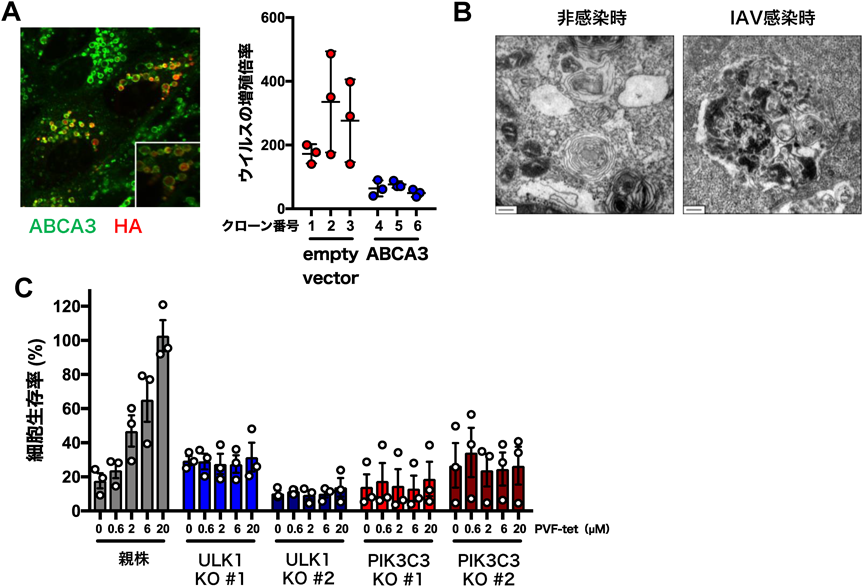
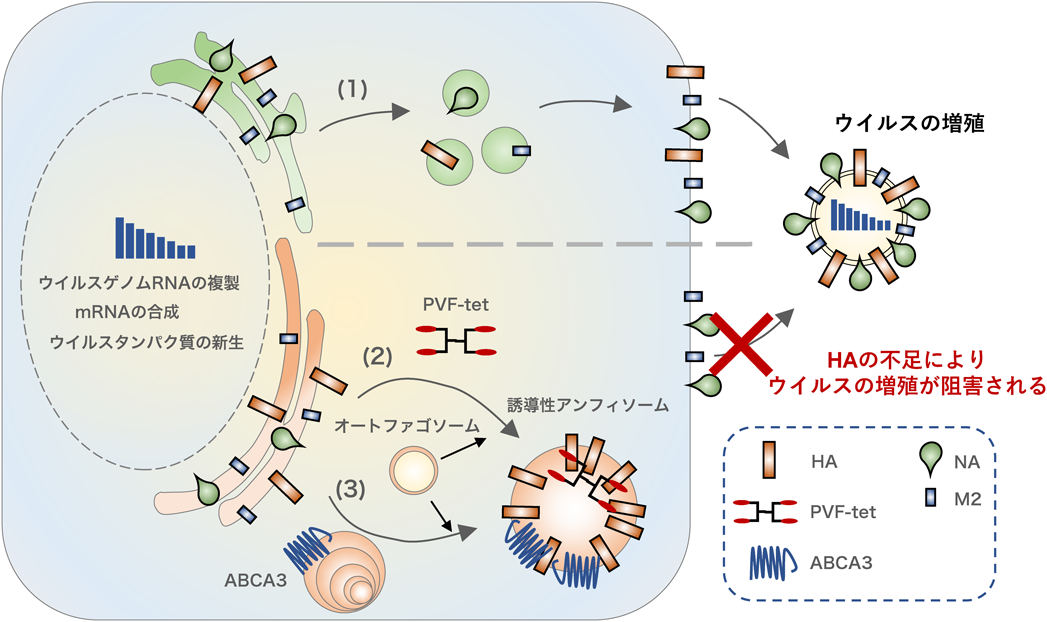
誘導性アンフィソームはインフルエンザウイルス感染防御に働くThe inducible amphisome defends against influenza A virus infection
1 東京大学大学院薬学系研究科衛生化学教室 特任助教Department of Health Chemistry, Graduate School of Pharmaceutical Sciences, The University of Tokyo ◇ 〒113–0033 東京都文京区本郷7–3–1 ◇ 7–3–1 Hongo, Bunkyo-ku, Tokyo 113–0033, Japan
2 同志社大学大学院生命医科学研究科分子生命化学 教授Faculty of Life and Medical Sciences, Doshisha University ◇ 〒610–0321 京都府京田辺市多々羅都谷1–3 ◇ 1–3 Miyakotani, Tatara, Kyotanabe, Kyoto 610–0394, Japan
発行日:2021年4月25日Published: April 25, 2021