piRNAは,トランスポゾンが内包されているゲノム中の「piRNAクラスター領域」から転写された長鎖RNA(このpiRNA前駆体もlncRNAの一種といえる)やトランスポゾン自身からプロセッシングされる小分子RNAである1–3).PiwiやMiwi2は,細胞質で生成された成熟型piRNAと複合体を形成し,核内で標的トランスポゾンの新生RNAに結合することで,抑制性ヒストン修飾であるヒストンH3の9番目のリシン残基に対するトリメチル化修飾(H3K9me3)の付与やDNAのメチル化を介して転写を抑制する.piRNAクラスターの発現様式やpiRISCの生合成についても近年さまざまな知見が蓄積されているが,ここではpiRISCによる転写抑制機構を中心に概説する.また,生合成が転写制御と協調して起こる分裂酵母siRNAに関しても,piRNAによる制御と比較しながら紹介する.
1)ショウジョウバエpiRNAによる転写サイレンシング
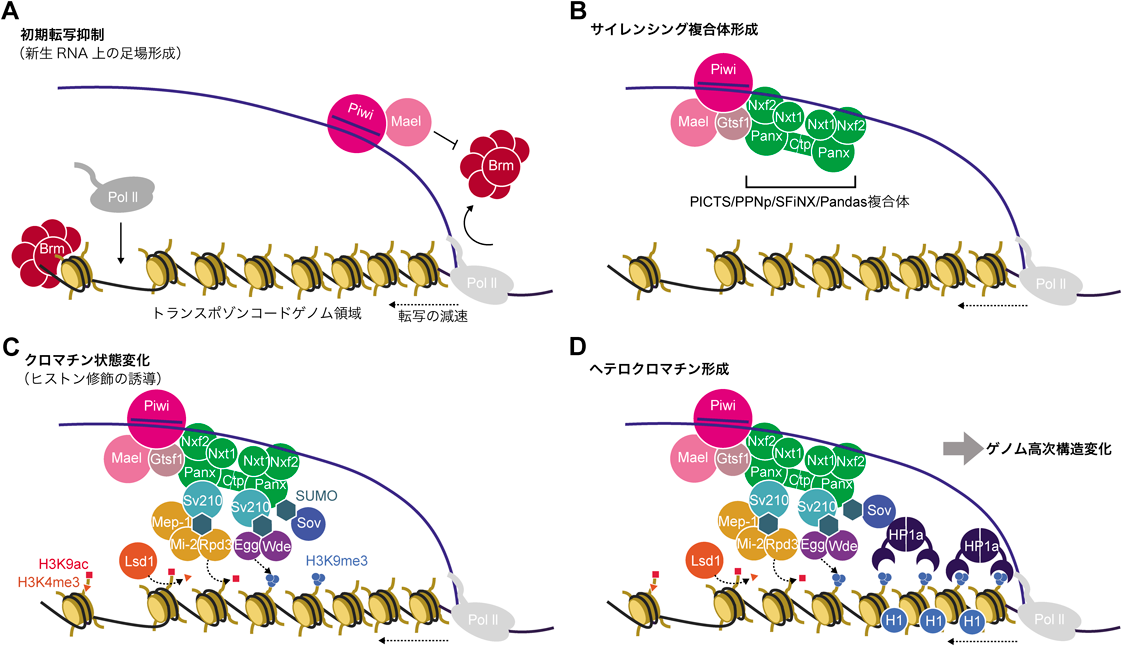
転写抑制複合体を新生RNA上に形成するpiRISCは,まず標的RNAに結合した直後に転写速度を低減されることで,安定的な足場を形成する.このプロセスに,Maelstrom(Mael)が寄与していることが示された19).Maelは以前からトランスポゾンのサイレンシングに不可欠であることが示されていたものの7, 20, 21),Mael欠損条件下でpiRNAを介した制御の際に付与されるH3K9me3の量にほとんど影響がないことから,MaelがどのようにpiRNAの転写抑制に関与しているかは不明であった7).しかしながら,近年の培養細胞系を用いた解析により,Maelは核内のPiwiと複合体を形成し,この複合体はさらにクロマチンリモデラーSWI/SNF複合体のコアユニットであるBrahma(Brm)に結合することが示された19).SWI/SNFはプロモーター領域周辺のクロマチン構造を弛緩させ,Pol IIを介した転写を促進することが知られている22).このことから,PiwiはMaelとともに標的トランスポゾン周辺のSWI/SNF複合体を阻害することで,転写速度を負に制御し,サイレンシング複合体形成のための足場を確立するというモデルが提唱された(図2A).
トランスポゾンの新生RNAに足場を形成したpiRISCは,次にサイレンシング複合体を呼び込むことで転写制御を開始する.サイレンシング複合体である,PICTS/PPNp/SFiNX/Pandasは,Piwiと相互作用するPanoramix/Silencio(Panx),Nxf2, Nxt1からなる23–26).λN-BoxBシステムを用いて人工的に任意のタンパク質をレポーターに係留する実験系により,PanxをレポーターRNAに係留すると,H3K9me3の蓄積とレポーターの転写制御が観察された27, 28).さらにこの制御は,Piwi非存在下でも確認されたことから,PanxがPiwiの下流で遺伝子サイレンシングを誘導することが示された.また,RNA核外輸送タンパク質ファミリーの一員であるNxf2は,幅広いmRNAの核外輸送因子として知られているNxf1とは異なり,核外輸送能を持たない23, 26).その一方でRNA結合ドメインは機能的であり,piRNAの標的トランスポゾンの新生RNAとpiRISCを安定的に結合させていることが示された25, 26).Nxt1/p15はもともとNxf1の補因子として報告されていたが29),これがNxf2との相互作用を介してpiRNAの転写抑制にも必須である.さらにこれら複合体構成タンパク質は,相互作用を通して互いを安定化させている23–26)(図2B).
近年,PICTS/PPNp/SFiNX/Pandas複合体の新たな相互作用因子として,Cutup/LC8(Ctp)が同定された30, 31).Ctpはユビキタスに発現し,二量体を形成することで安定な複合体を形成する32).Piwi-piRNA経路においても,Ctpを介してPICTS/PPNp/SFiNX/Pandas複合体が重合体を形成している.Gametocyte-specific factor/Asterix(Gtsf1)もまた,Piwiと相互作用する核タンパク質である33–35).詳細な作用機序は不明だが,Gtsf1欠損培養細胞では,Piwi標的トランスポゾンのH3K9me3修飾は低下し,転写の脱抑制が起こる.さらに,Gtsf1はpiRISCおよびPICTS/PPNp/SFiNX/Pandas複合体と相互作用していることから,piRISCの転写抑制ステップに必須であると考えられる19)(図2B).
続いてサイレンシング複合体は,転写を抑制するための酵素群をリクルートする.ショウジョウバエでは三つのH3K9特異的ヒストンメチル化酵素,Su(var)3-9, G9a, Eggless/SetDB1(Egg)が知られているが,これらのうちPiwiを介した転写サイレンシングで機能するのはEggである27).EggはUbc2によってユビキチン化されることでメチル化活性を示し,さらに補因子であるWindei(Wde)が結合することでクロマチン上に安定的に結合する36).また,H3K4脱メチル化酵素Lsd1が活性型ヒストン修飾であるH3K4のメチル化の除去に寄与する28).piRNA標的トランスポゾン領域は,Eggによるメチル化を受ける前に脱アセチル化される.この脱アセチル化に関与しているのが,Mi-2, Mep-1, Rpd3の複合体である37).ATP依存性のクロマチンリモデラーであるMi-2およびRpd3は,ショウジョウバエのNuRD複合体の一員である38).Mi-2については,ショウジョウバエMec複合体にも含まれる一方で,Rpd3は含まれない39).したがって,ここで形成されるMi-2, Mep-1, Rpd3を含む複合体は,Piwi-piRNAを介した転写抑制経路に特有のものであると考えられる37).さらに,E3リガーゼであるSu(var)2-10(Sv210)による,Egg, Wde, Mi-2, Mep-1のSUMO化がサイレンシング複合体との相互作用に重要な役割を果たす可能性が示唆されている37, 40).Sv210を強制的にレポーターに係留した際に転写抑制を引き起こすためには,Wde-EggとSv210によるSUMO化活性の両方が必須であった40).このことから,Sv210は自身を含む近接タンパク質をSUMO化し,Egg, Wde, Mi-2, Mep-1をリクルートするための足場を提供している可能性が考えられる.近年,PanxもSUMO化を介してzinc finger proteinであるSmall ovary(Sov)をリクルートし,ヘテロクロマチン形成を誘導することが報告された41).多くのpiRNA経路関連因子がSUMO相互作用モチーフを持つことから,SUMO化が転写制御複合体形成の鍵となっていると考えられ,今後のさらなる解析がまたれる(図2C).
上記のとおり抑制性ヒストン修飾が付与されたpiRNA標的トランスポゾン領域に対して,ヘテロクロマチンタンパク質1a(HP1a)が結合することでヘテロクロマチンが維持される42–44).また,リンカーヒストンであるヒストンH1は,Piwiを介してトランスポゾン領域に安定的に結合し,piRNA標的トランスポゾン領域のクロマチンアクセシビリティを低下させる45).さらに,ラミン相互作用領域の網羅的解析法であるLamin DamID解析を用いた結果から,piRISCによる制御が標的トランスポゾンコードゲノム領域を核膜付近に係留すること,およびゲノム高次構造の網羅的解析法であるHiC解析から近隣ゲノム領域間の相互作用を増強することで,安定的なヘテロクロマチン構造が維持されていることが明らかとなった46).以上のように,ショウジョウバエでは,さまざまな因子とともにPIWI-piRNAが段階的かつ綿密なトランスポゾン抑制ステップを踏んで転写の抑制状態を維持していることが明らかになっている(図2D).
2)マウスpiRNAによる転写サイレンシング
ショウジョウバエのPiwiは卵巣と精巣の両方で機能するのに対して,マウスのPiwiホモログであるMiwi2(Piwil4)の機能は精巣特異的である47).マウスのほかのPIWIタンパク質であるMiwi(Piwil1)とMili(Piwil2)もほぼ精巣特異的に発現するが,ショウジョウバエのAubとAGO3と同様に細胞質で転写産物を切断・分解することでトランスポゾンの発現を抑制する5, 48–51).実際に,各マウスPIWIタンパク質について,スライサー活性の必要性を検討した結果,Miliのスライサー活性変異マウスでは,胎生期の精巣におけるトランスポゾン由来のpiRNAの増幅が行われておらず,Mili欠損と同様に精子形成不全による不妊を引き起こした.その一方で,同じ胎生期の精巣に発現するMiwi2のスライサー活性変異マウスでは正常にpiRNAが増幅し,精子形成が行われていた.このことからも,Miwi2によるサイレンシングはRNAの分解を介したものではなく,ショウジョウバエPiwiと同様にエピジェネティックな転写抑制機構であるといえる5).すべてのマウスPIWIタンパク質は精子形成に必要だが,精子形成中の発現時期は異なる.Miwiはパキテン期から円形精子細胞まで発現し49, 51),Miwi2は精原幹細胞(SSCs)の前駆細胞に特異的であり47),Miliはこれらの細胞すべてで発現する48, 50, 51).
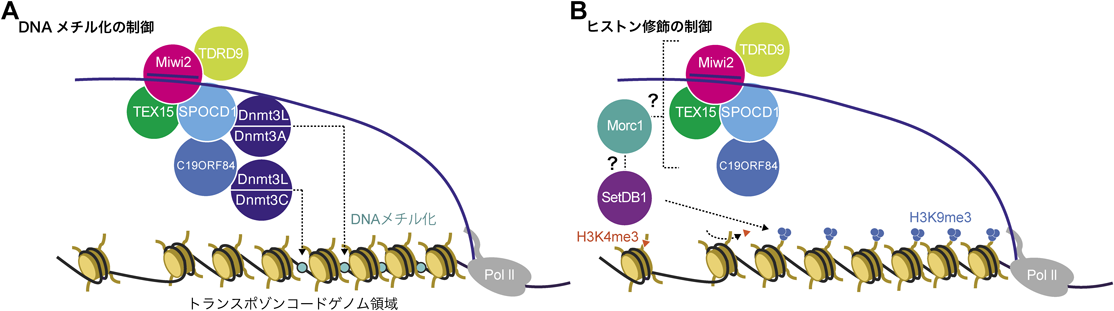
哺乳類の生殖サイクルでは,受精卵と始原生殖細胞(PGC)の2回にわたり初期化(リプログラミング)が起こり,エピジェネティック修飾が消去される52, 53).生殖細胞では,このリプログラミングの後,精子形成を適切に進めるために,ゲノム全体でde novo DNAメチル化が起こる41, 54–58).de novo DNAメチル化が起こる雄性生殖細胞はゴノサイトと呼ばれ,MiliとMiwi2を発現している.Mili-piRISCが細胞質で標的RNAを切断し,その切断産物がpiRNAとしてMiwi2に結合する5).その後,Miwi2-piRISCは,DNAメチル化と抑制性ヒストン修飾を介してトランスポゾンの転写を抑制する47, 59).マウスは四つのDnmt3メンバー,Dnmt3A, Dnmt3B, Dnmt3L, Dnmt3Cを発現しており,これらすべてが正常な精子形成に必要である60–65).これらのうち,齧歯類特異的なDnmt3Cは生殖細胞特異的に発現していることが知られており,その枯渇はトランスポゾンプロモーター領域のDNA脱メチル化を特異的に引き起こす60).このことから,Dnmt3CがマウスPIWI-piRNA経路でDNAメチル化を引き起こしていると考えられる(図3A).
Miwi2は,ゴノサイト期に複数の因子と相互作用することが報告されており,TDRD9もその一員である66, 67).TDRD9はTudorドメインを含むDExH-boxヘリカーゼ/ATPaseで,胚と成体の両方の精巣で発現している.tdrd9変異個体では,トランスポゾンのDNAメチル化に異常が確認された一方で,piRNAの産生量には影響がみられなかった.このことから,TDRD9は特にトランスポゾンの転写抑制の際に重要な機能を持つことが予想されるが,詳細なメカニズムに関しては明らかになっていない.さらに近年,Miwi2の新たな相互作用因子として,SPOCD1とTEX15が同定された68–70).SPOCD1は,TFIISとSPOCドメインを持ち,TEX15はDUF3715と二つのTEX15ドメインを持つ.SPOCD1およびTEX15の欠損個体では減数分裂が早期に停止し,完全な不妊となる.この変異体ではトランスポゾン上のDNAメチル化レベルが低下し,トランスポゾンの発現が脱抑制される.SPOCD1と相互作用する因子には,NuRD複合体とBAF複合体の構成要素が含まれるが,これらの複合体がpiRNAを介した転写制御でどのような機能的役割を果たすかについては未解明である70).さらにSPOCD1は,C19ORF84との相互作用を介してDnmt3Cをリクルートすることが示された71).このことから,SPOCD1が誘導するDnmt3Cの作用により,Miwi2-piRNAはトランスポゾン領域にDNAメチル化を付与するというモデルが考えられる.加えて,C19ORF84のヒトホモログにおける変異がヒトの不妊症患者で確認されたことから,このトランスポゾン抑制機構がヒトでも保存され,かつ重要な生理的意義を持つ可能性が示唆された71)(図3A).
Morc1はMorcファミリーに属し,その枯渇は雄性不妊の原因となることが古くから知られていた72, 73).Morc1は胚盤胞や生殖細胞で高発現している一方で,分化した細胞では発現しておらず,morc1変異マウスでは,トランスポゾンの低メチル化と発現上昇が観察されるが,piRNAの産生量には影響しない74).その一方で,Morc1の影響を受けるゲノム領域がpiRNA標的トランスポゾン領域に限られないことから,piRNA経路における機能については不明であった.近年,ゴノサイトの解析を通して,Morc1が生殖細胞におけるMiwi2標的トランスポゾンにH3K9me3修飾を付与する上で重要な役割を果たしていることが示唆された75).その一方で,Morc1とMiwi2やSetDB1との相互作用は観察されていないことから,どのようにMorc1がMiwi標的トランスポゾン領域を認識するかについては,今後の課題となっている(図3B).
マウスPIWI-piRNA経路においては,DNAのメチル化が起こらないショウジョウバエと異なり,de novo DNAメチル化を介した制御と抑制性ヒストン修飾が協調して機能していると考えられる.精子形成と卵形成の初期発生段階では,de novo DNAメチル化の前に,SetDB1がトランスポゾン遺伝子座のH3K9をメチル化することが,その後の適切なDNAメチル化に重要であるといわれている76).また,PIWI-piRNA関連因子の変異個体について,脱抑制は起こるもののDNAメチル化状態に大きな変動がない種類のトランスポゾンも観察されている70, 71).このことから,トランスポゾンの種類によって,転写抑制の際のDNAメチル化とヒストン修飾の依存度合が異なる可能性もある.DNAメチル化の制御と抑制性ヒストン修飾の付与がPIWI-piRNA制御で互いにどのような関連性にあるかについては,今後明らかになっていくことが期待される(図3).
3)分裂酵母小分子RNAによるヘテロクロマチン形成
ショウジョウバエやマウスの核内でPIWI-piRNA経路がトランスポゾンの転写を抑制するメカニズムに加え,分裂酵母ではAgo1とsiRNAの複合体を介した構成的ヘテロクロマチン形成がセントロメア周辺領域やサブテロメア領域で起こる.それぞれの遺伝子座にはリピート配列があり,この領域からsiRNAが産生され,複数の相互作用因子と協調して染色体凝縮を引き起こす77–79).さらにこの染色体凝縮は隣接領域にも広がり,遺伝子座を完全に覆うようになる80, 81).このsiRNA依存的な構成的ヘテロクロマチン形成は,組換えや細胞分裂時の染色体分配など,染色体の正常な機能に重要な役割を果たす82).
リピート配列は構成的ヘテロクロマチン領域内に存在するが,siRNAはRNAを鋳型として産生されるため,転写されることが必須となる83).転写された新生一本鎖RNAは,RNA-directed RNA polymerase complex(RDRC)の一員であるRdp1によって二本鎖RNA(dsRNA)に逆転写される80, 84).RDRCにはRdp1のほかに,ヘリカーゼであるHrr1と,ポリ(A)ポリメラーゼファミリーメンバーであるCid12が含まれる80).dsRNAは,ヒトDicerの分裂酵母オルソログであるDcr1によって,21塩基の二本鎖siRNAにプロセシングされる85, 86).二本鎖のsiRNAはAgo1と複合体を形成し,siRNAとして働かない方の鎖はAgo1のスライサー活性によって切断され分解される87).Ago1とsiRNAはさらにChp1とTas3とともにRNA-induced transcriptional silencing(RITS)複合体を形成し,サイレンシングのプラットフォームとして機能する78, 88).
RITS複合体は標的のリピート新生RNAを認識し,Clr4メチル化酵素複合体(CLRC)をリクルートする89–91).Clr4は,哺乳類のSuv39hやショウジョウバエのSu(var)3-9のホモログのH3K9メチル化酵素である92–94).CLRCはH3K9me3を付与し,これにRITS複合体内のChp1がクロモドメインを介して結合し,RITSのヘテロクロマチンへの結合を安定化することで転写を抑制する81, 95–97).
分裂酵母のRITSが引き起こすサイレンシングと,piRISCが引き起こす転写サイレンシングは同様にヘテロクロマチン形成を介するなど,多くの共通点がみられる.その一方で,分裂酵母では,siRNAは自身の前駆体が産生される遺伝子座を標的とすることで,シスにヘテロクロマチン形成を誘導する.それに対してPIWI-piRNAはpiRNAクラスターから産生され,配列的な相補性をもって標的をトランスに認識し転写抑制するといった違いもみられる.分裂酵母のsiRNAは構成的ヘテロクロマチン領域を安定的に維持しているのに対して,piRNAはホストにとって有害な転写活性を持つトランスポゾンの抑制に機能する,といったサイレンシング機構が果たす役割の違いもある.ゲノム中に散在するトランスポゾンを効率的に抑制するためには,トランスに作用できることが効率的であることも想像できる.
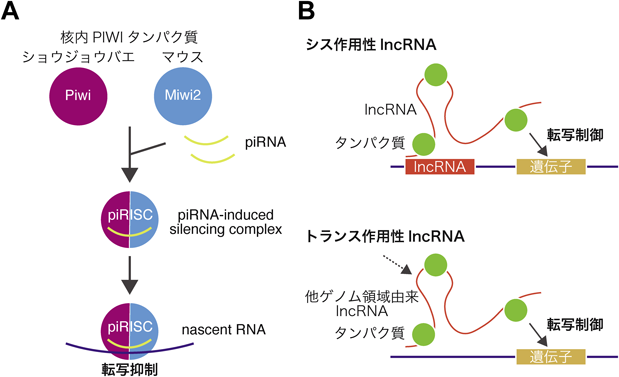
現在ではさまざまなlncRNAがエピジェネティックな機構を介して転写抑制に関与することが知られているが,その先鞭となったのが遺伝子量補償を制御するlncRNAである.遺伝子量補償とは,生物が両性における遺伝子発現を均等に保つメカニズムである.ここでは,哺乳類XISTならびにショウジョウバエroXによる染色体レベルの転写調節について概説する.
1)哺乳類XISTによるX染色体遺伝子量補正
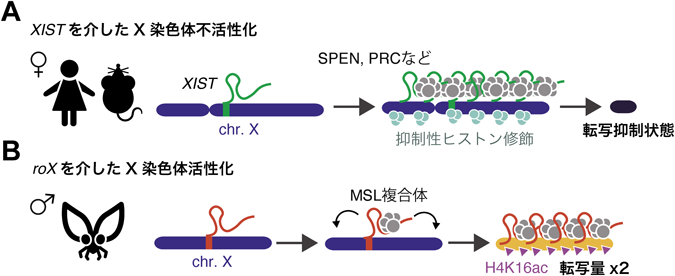
哺乳類雌で機能するlncRNAのXISTは,X染色体の不活性化を担っている.胚発生の過程で,XIST分子は2本のX染色体の一方に広がり,その遺伝子の大部分をサイレンシングする98).XISTは,X染色体以外の染色体から異所的に発現させた場合でも,XISTを発現した染色体領域をサイレンシングすることができる99).このことから,XISTの発現自体が制御対象となっている染色体領域を決定していると考えられる.サイレンシングを引き起こしているのは,XISTがリクルートするさまざまなタンパク質である100–103).XISTは転写抑制因子として知られるSPENやポリコーム複合体など種々のエピジェネティック制御因子を制御対象X染色体に呼び込み,抑制性ヒストン修飾の付与を介して遺伝子発現を抑制すると考えられているが,この抑制複合体については未解明の部分も大きい.さらに,マウス胚性幹細胞を用いた研究から,XISTがX染色体を迅速に覆うために,ゲノムの三次元構造を利用していることが示唆されている.これにより,XISTは空間的に近接したゲノム領域から異なる遺伝子座へと拡散し,抑制性ヒストン修飾を誘導する因子との相互作用を通して標的のゲノム領域を抑制する104).XISTによる転写抑制は,一度開始されるとXISTが存在しない場合でも持続することが示されていることから100–103),XISTはエピゲノム状態制御のイニシエーターとしての役割を果たしているといえる(図4A).
2)ショウジョウバエroXによるX染色体遺伝子量補正
ショウジョウバエの性決定も哺乳類と同様に,XXが雌,XYが雄であることが知られているが,哺乳類とは異なり雄のX染色体からの転写量を倍に増幅することで,雌雄間の染色体数に起因する発現量の差を補っている105–107).ショウジョウバエでは,二つのroX lncRNAであるroX1とroX2が五つのMSL(male-specific lethal)タンパク質(MLE, MSL1/2/3, MFO)とともにMSL複合体を形成する108, 109).この複合体が,活性型ヒストン修飾であるH4K16acを付与することによって,単一の雄X染色体上の遺伝子発現を促進し,雌における二つのX染色体由来の発現量とのバランスを保つ.MSL1はRNAと結合するMSL3を含むほかのタンパク質と相互作用し,MSL1/2はX染色体上の高親和性部位として知られる結合部位を認識する110).さらに,MOFはH4K16acを誘導し,活性型ヒストン修飾を広げる108).roX1が欠損するとX染色体遺伝子の発現が減少し,roX2が欠損するとMSLに依存しない遺伝子発現が起こる111).このようにroX1とroX2がそれぞれ独立して機能している点も興味深い(図4B).
哺乳類ではXISTが主にX染色体のシスに広がって機能するが,ショウジョウバエのroX1/2はトランスで機能するというユニークな特徴を持っており112),またXISTは抑制に働く一方でroXは活性化に働くという点でも異なる.これは,遺伝子量補償といった共通の現象の制御を担うlncRNAの機能的多様性を示している.また,X染色体全体といった広範囲の制御には,長鎖RNAの分子量がタンパク質と比較して大きいという性質が適しているとも考えられる.lncRNAの作用機構は多岐にわたることから,今後もユニークかつ生体に必要不可欠な機能が発見される可能性にも期待したい.